15 методов раскрутки сайта, актуальные в 2020 году — Маркетинг на vc.ru
Методы раскрутки сайтов
72 616 просмотров
Раскрутка сайта – это комплекс действий, направленных на привлечение максимального количества пользователей из сети Интернет на ваш веб-сайт.
Под понятием “раскрутка сайта” многие понимают абсолютно разные вещи, от SEO (поисковая оптимизация сайта) до рекламы и, даже, брендинга.
Отсюда, кстати, происходит полное непонимание, когда речь заходит о ценах:
— “Сколько стоит раскрутка сайта у вас?”
— “В среднем 45-90 тыс. руб”
— “А, нет, мне вот за углом за 5 тыс. предлагают…”
 п. И никому не приходит в голову придти в салон Мерседес и сказать, — “а, за углом за 5 тыс. можно тоже машину купить”.
п. И никому не приходит в голову придти в салон Мерседес и сказать, — “а, за углом за 5 тыс. можно тоже машину купить”.В данной статье я хочу разрушить множество мифов и показать, а что же в принципе входит в понятие “раскрутка сайта”, какие механики для этого существуют, какие инструменты и методы, а отсюда и вопрос с ценой решится сам собой.
Если говорить все же про определение, то, наверное, наиболее правильным будет отталкиваться от цели этого процесса, а цель — привлечение целевых посетителей с целью совершения ими определенного действия на нем, например, покупка, звонок, регистрация, загрузка прайс-листа и т.п.
1) Поисковая оптимизация (SEO)
Поисковая оптимизация (SEO) — оптимизация сайта с целью получения высоких рангов в поисковой системе по запросам, отвечающим тематике сайта. Существует 5 основных стратегий работы с теми или иными видами поисковых запросов:
Существует 5 основных стратегий работы с теми или иными видами поисковых запросов:
- брендинговые запросы (содержащие название бренда вашей компании)
- брендинговые запросы конкурентов (содержащие название бренда конкурента)
- горячие или продающие запросы (например, “купить ноутбук MacBook Pro”)
- запросы, связанные с отзывами и мнениями (например, “macbook air i5 отзывы”)
- информационные запросы (например, “что такое раскрутка сайтов”)
Сервис подбора ключевых фраз от Яндекс
Рекомендация: Поисковую оптимизацию (SEO) имеет смысл использовать, если:
- Продукт и услуга уже давно присутствуют на рынке.

- Не все места в ТОП-10 поисковых систем заняты сайта-агрегаторами, маркетплейсами, досками объявлений и информационными или новостными ресурсами. В лидерах есть сайты ваших прямых конкурентов.
- Ассортимент товаров вашего сайта соизмерим с сайтами-конкурентами.
- Предоставляемые услуги или товар имеют соизмеримое качество и соизмеримую цену с конкурентами. Хорошо, если есть выработанное уникальное торговое предложение (УТП).
- Есть достаточный бюджет на доработку, наполнение сайта минимум на срок от 4 до 12 месяцев проведения работ по SEO.

2) Контекстная реклама
Контекстная реклама — показ рекламных объявлений пользователям, заинтересованным в них в данный конкретный момент поиска;
- Яндекс. Директ
- Google Ads
 Директ
ДиректРекомендация: Если вы только планируете запускать свои первые рекламные кампании контекстной рекламы, то рекомендую обратить внимание на
3) Контекстно-медийные сети
- GSP — реклама в почте GMail.com
- КМС Google
- Рекламная сеть Яндекса (РСЯ)
 Например, «Одежда, приносящая деньги и удачу».
Например, «Одежда, приносящая деньги и удачу».Картинки: Используйте яркие картинки, цепляющий call to action (Не покупайте натяжной потолок! Пока не ознакомитесь с нашим предложением).
4) Медийная реклама
Медийная реклама — показ рекламных баннеров на сайтах; Это больше относится к брендинговой рекламе, т.к. основная задача сводится не к конверсии на сайте, а к тому, чтобы примелькаться.
Рекомендация:
 руб и основная задача — конверсия.
руб и основная задача — конверсия.5) Отраслевые и прайс-агрегаторы
Отраслевые и прайс-агрегаторы— представляют собой площадки с каталогами товаров от разных компаний, например:
- Яндекс.Маркет (Яндекс)
- Prise.ru (Рамблер)
- Beru.ru (Яндекс и Сбербанк)
- Goods.ru (МВидео)
- Товары Mail.ru (Mail.ru)
Рекомендации: Прайс-агрегаторы идеально подходят для продвижения интернет-магазинов. Стоит помнить, что для увеличения объема продаж и спроса необходимо всегда располагать привлекательными ценами, т.к. ранжирование зачастую идет с использованием цены и качества обслуживания.
Стоит помнить, что для увеличения объема продаж и спроса необходимо всегда располагать привлекательными ценами, т.к. ранжирование зачастую идет с использованием цены и качества обслуживания.
6) Таргетированная реклама в социальных сетях
Таргетированная реклама в социальных сетях позволяет показывать рекламные объявления в соответствии с демографическими характеристиками и персональными интересами пользователя;
- Vkontakte
- Одноклассники (MyTarget)
- YouTube (Google Ads)
Функционал очень обширный, я записал для вас полное руководство по всем функциям данного инструмента, доступ открыт только по ссылке на YouTube (промо-код на бесплатное использование инструмента — semcomplex)
7) Партнерские программы
Партнерские программы (англ. Affiliate program) или партнёрка — форма делового сотрудничества между продавцом и партнёрами, при продаже какого-либо товара или предоставления услуг с оплатой в % с продажи.
Affiliate program) или партнёрка — форма делового сотрудничества между продавцом и партнёрами, при продаже какого-либо товара или предоставления услуг с оплатой в % с продажи.
- justclick.ru
- autoweboffice.com
- e-autopay.com
- onwiz.ru
- glopart.ru
- goodly.pro
- spoonpay.com
- autooffice24.ru
- orderbro.ru
Рекомендация: При выборе партнерской программы придерживайтесь следующих 5 пунктов: удобство регистрации для пользователей, удобство зоны для адвертов, разнообразие вариантов для выплаты комиссионных, полная статистика, быстрая поддержка. Мы нашим клиентам часто устанавливаем партнерскую программу от сервиса justclick.
Мы нашим клиентам часто устанавливаем партнерскую программу от сервиса justclick.
8) Видеореклама и мобильная реклама
Видеореклама и мобильная реклама пока используется компаниями в качестве тестового канала, но постепенно набирает обороты, особенно мобильная реклама, темпы роста которой, по различным оценкам, превышают 40 % в год;
- YouTube (через сервис Google Ads)
- Социальные сети
- Мобильные сети
- Сети таргетированной рекламы
- Онлайн-кинотеатры
Рекомендация: Для создания видео-контента можете использовать сервис supa. ru
ru
9) Email-маркетинг
Email-маркетинг — полезные рассылки по электронной почте пользователям, давшим на это добровольное согласие;
- Кросс-рассылки по партнерским базам
- Информационные периодические рассылки
- Триггерные рассылки
Рекомендация: Для рассылок по партнерским базам с целевой аудиторией можете воспользоваться сервисом subscribe.ru или bazaarmail.com
10) Контент-маркетинг
Контент-маркетинг — привлечение пользователей за счёт публикации полезного контента. Это тот контент, который решает проблему читателя. Контент можете писать самостоятельно, либо воспользоваться помощью копирайтеров. Биржи копирайта:
Это тот контент, который решает проблему читателя. Контент можете писать самостоятельно, либо воспользоваться помощью копирайтеров. Биржи копирайта:
- eTxt.ru
- advego.com
- text.ru
Рекомендация: Если пользуетесь биржами-копирайтинга, обязательно создавайте white-листы, т.е. белые списки тех хороших копирайтеров, чьими текстами вы удовлетворены по той или иной тематике.
11) Social Media Marketing (SMM)
Social Media Marketing (SMM) — создание лояльной аудитории в социальных сетях благодаря публикации полезной информации и общению с потенциальными потребителями.
Рекомендация: используйте сервисы автоматизации при работе с SMM, например, сервис Super SMM, видео-презентацию данного сервиса доступна по ссылке выше.
12) Выдача в новостных агрегаторах
- Яндекс.Новости
- Google News
Рекомендация: Для того, чтобы воспользоваться возможностью использования новостных агрегаторов, необходимо внедрить на своем ресурсе специальный новостной блок, либо вести блог с уникальным и полезным новостным контентом.
13) Работа с отзывами и упоминаниями
- iqbuzz.pro
- babkee.ru
- google alerts
Рекомендация: Зарегистрируйтесь на бесплатном сервисе от Google, а именно, Google Alerts (google.ru/alerts) и отслеживайте все новые статьи, где встречается адрес вашего сайта, название вашего бренда, ваша ФИО, ФИО руководителя и генерального директора.
14) Ретаргетинг/ремаркетинг
- Поисковый
- Динамический
- CRM-ретаргетинг (загрузка Email и телефонов)
Рекомендация: Если есть база телефонов, то лучше использовать ее. База Е-майлов тоже работает, но, практика показывает, по телефонам все рекламные сети ищут гораздо лучше.
База Е-майлов тоже работает, но, практика показывает, по телефонам все рекламные сети ищут гораздо лучше.
15) Мобильная реклама
- Собственное мобильное приложение (пуш-уведомления)
- Баннеры и фуллскрины
- Интеграция и мобильные игры
Рекомендация: Не обязательно создавать собственное мобильное приложение с нуля, можете воспользоваться конструктором мобильных приложений. Детальный разбор разных конструкторов доступен на VC
Этот список нельзя назвать исчерпывающим перечнем всех возможных инструментов раскрутки, однако это наиболее часто используемые методы.
Буду благодарен за ваше мнение в комментариях — какие методы раскрутки сайта используете Вы?
Продвижение сайтов и раскрутка сайтов в Яндексе и Google
Продвижение сайта в поисковых системах остается наиболее эффективным способом привлечения новых клиентов и роста продаж в интернете. Многие специалисты сходятся во мнении, что органический трафик из результатов поиска имеет наибольшую конверсию. Раскрутка сайтов в Яндексе и Google — это предоставление персонализированной информации по запросу пользователя, которая необходима ему здесь и сейчас. Именно поэтому трафик с поиска приносит «горячих» клиентов, а вам — гарантированные продажи.
Мы эффективно сочетаем контекстную рекламу и SEO-продвижение сайтов
Для обеспечения комплексного продвижения вашего бизнеса в интернете агентство Registratura. ru совместно с компанией iSEO предлагает интегрированный продукт, сочетающий контекстную рекламу и поисковую оптимизацию в рамках единой digital-стратегии. В зависимости от тематики и технического состояния сайта продвижение в поиске часто требует больше времени, в отличие от контекстной рекламы, где результат появляется моментально. Грамотное сочетание этих способов продвижения в интернете позволяет добиться максимальной эффективности каждого канала. Подключение контекстной рекламы на момент начала продвижения сайта или при сезонном падении спроса в поиске позволяет не потерять объемы продаж, а более оперативный эффект от рекламы дает возможность оценить конверсию ключевых слов и скорректировать стратегию по оптимизации и продвижению сайта.
ru совместно с компанией iSEO предлагает интегрированный продукт, сочетающий контекстную рекламу и поисковую оптимизацию в рамках единой digital-стратегии. В зависимости от тематики и технического состояния сайта продвижение в поиске часто требует больше времени, в отличие от контекстной рекламы, где результат появляется моментально. Грамотное сочетание этих способов продвижения в интернете позволяет добиться максимальной эффективности каждого канала. Подключение контекстной рекламы на момент начала продвижения сайта или при сезонном падении спроса в поиске позволяет не потерять объемы продаж, а более оперативный эффект от рекламы дает возможность оценить конверсию ключевых слов и скорректировать стратегию по оптимизации и продвижению сайта.
Мы применяем высокие стандарты продвижения сайтов
Поисковые алгоритмы меняются, но неизменным остаются стандарты качества нашей работы — адаптация сайта под алгоритмы поисковых систем, тенденций рынка и ожидания пользователей. Наша цель — достижение бизнес-задач клиента за счет привлечения качественного конверсионного органического трафика. Для этого мы глубоко погружаемся в каждый проект, проводим тщательный анализ рынка и конкурентов, проверяем сайт на наличие ошибок, работаем над повышением конверсии. Разрабатываем стратегию, при которой раскрутка сайта будет максимально быстрой и эффективной. В этом нам помогает многолетний опыт работы с проектами различных масштабов и тематик и глубокое знание алгоритмов поисковых систем.
Для этого мы глубоко погружаемся в каждый проект, проводим тщательный анализ рынка и конкурентов, проверяем сайт на наличие ошибок, работаем над повышением конверсии. Разрабатываем стратегию, при которой раскрутка сайта будет максимально быстрой и эффективной. В этом нам помогает многолетний опыт работы с проектами различных масштабов и тематик и глубокое знание алгоритмов поисковых систем.
Мы постоянно развиваем свою экспертизу в области SEO и оказываем услуги по следующим направлениям:
Мы решаем задачи каждого клиента в персональном порядке и гибко подходим к условиям заключения договоров.
Разделяем риски и несем финансовую ответственность за результат
Мы работаем с бизнесами различных масштабов и готовы предложить выгодные условия каждому нашему клиенту. Помимо типовых схем продвижения сайтов в ТОП-10 и по трафику, мы можем согласовать с вами индивидуальную модель сотрудничества и закрепить важные для вас KPI в договоре.
Мы разделяем финансовые риски со своими клиентами и готовы работать по постоплате, что является лучшей гарантией для вас. Расчеты за продвижение сайта происходят по факту достигнутого результата. В конце месяца вы получаете подробный отчет, в котором зафиксированы ежедневные результаты по KPI, на основе этих данных и формируется счет за наши услуги.
Расчеты за продвижение сайта происходят по факту достигнутого результата. В конце месяца вы получаете подробный отчет, в котором зафиксированы ежедневные результаты по KPI, на основе этих данных и формируется счет за наши услуги.
Мы придерживаемся принципа справедливой цены — стоимость работ по каждому проекту мы оцениваем на основе реальных трудозатрат, необходимых для достижения желаемого результат. Мы не готовы демпинговать цены ради заключения договора, так как для нас на первом месте стоит качество оказываемых услуг, а не количество проектов. Если имеющийся бюджет не позволяет получить желаемый результат, мы открыто говорим об этом, предлагая вам альтернативные методы продвижения вашего бизнеса.
Мы зарекомендовали себя как профессионалы в области SEO
Мы специализируемся исключительно на поисковом продвижении сайтов, все наши сотрудники являются настоящими экспертами в области SEO и имеют многолетний опыт работы с проектами различных масштабов и тематик. Мы в курсе всех изменений, происходящих в алгоритмах поисковых систем, и умеет быстро адаптировать стратегию продвижения под новые условия без потери результата. Мы постоянно повышаем наши знания и экспертизу, стремимся быть лучшими в отрасли и предлагаем своим клиентам только эффективные методы оптимизации сайта, которые мы формируем индивидуально под каждый проект.
Мы в курсе всех изменений, происходящих в алгоритмах поисковых систем, и умеет быстро адаптировать стратегию продвижения под новые условия без потери результата. Мы постоянно повышаем наши знания и экспертизу, стремимся быть лучшими в отрасли и предлагаем своим клиентам только эффективные методы оптимизации сайта, которые мы формируем индивидуально под каждый проект.
К нашему мнению прислушиваются, ежегодно мы являемся организаторами секций и докладчиками на крупнейших отраслевых мероприятиях: «РИФ+КИБ», RIW, eTarget и др.
Наши методы работы и успешные результаты помогли нам приобрести репутацию надежного партнера и эффективного подрядчика. 99% клиентов довольны нашей работой, ни один сайт нашего клиента не попал под фильтр поисковых систем, многие клиенты работают с нами на протяжении нескольких лет. Залог успеха заключается в отсутствие пустых обещаний, наши гарантии строятся на практическом опыте и алгоритмах работы поисковых систем.
В прозрачности нашей работы и качестве услуг убедились многие компании: Avito, L’Оreal, Volvo, KIA, «Ингосстрах», банка «Ренессанс Кредит», «Банка Хоум Кредит», «ЮниКредит Банк», Biglion, Аптека. ру, «Адамас», MEXX, ESET и др.
ру, «Адамас», MEXX, ESET и др.
Мы готовы предложить вам разработку и реализацию индивидуальной digital-стратегии по эффективному продвижению сайта в ТОП-10 и увеличению качественного трафика на сайт.
Звоните нам по телефону в Москве +7 (495) 221-68-19
Пишите на sales AT iseo DOT ru
Раскрутка и продвижение сайта цена за месяц в Москве, стоимость seo оптимизации
Можете ли Вы гарантировать, что сайт попадет в ТОП 10?
На первом этапе проводится поисковый аудит, после чего мы разрабатываем стратегию продвижения сайта с учетом особенностей Вашего бизнеса и региона, в который Вы ориентированы. Мы делаем все для роста позиций и видимости в сети, а также для увеличения звонков и заявок. Но Вы должны понимать, что попадание в ТОП выдачи зависит не только от нас, но и от поисковых систем, которые постоянно меняют алгоритмы и предъявляют новые требования к контенту.
Как много времени потребуется, чтобы попасть в ТОП 10?
В каждом случае сроки индивидуальны и зависят от текущего состояния сайтов, активности конкурентов, скорости и качества работ SEO специалистов. Первые результаты могут появиться спустя месяц с начала работ. Полностью оценить эффективность проведенных работ можно спустя 4 месяца.
Первые результаты могут появиться спустя месяц с начала работ. Полностью оценить эффективность проведенных работ можно спустя 4 месяца.
Чем отличаются ваши тарифы?
Мы предоставляем разные услуги, а также оказываем разный объем работ. Более подробную информацию по каждому тарифу можно получить на консультации. Наши специалисты помогут подобрать тариф, в соответствии с Вашими потребностями и возможностями.
Как выбрать оптимальный тариф?
Выбор в пользу того или иного пакета услуг зависит от разных параметров: тематики бизнеса, особенностей сайта, целей, которых Вы хотите добиться — рост трафика, увеличение количества заказов, вывод в ТОП, поддержание лидирующих позиций и т.д.
В зависимости от этих и параметров мы подбираем схемы и методики поискового продвижение сайта, поэтому и стоимость тарифов разная. Если Вы не можете определиться с нужным тарифом самостоятельно, наши специалисты проведут экспресс аудит Вашего сайта и подскажут, какой пакет услуг Вам необходим.
Как я могу оценить эффективность Вашей работы?
Успешность SEO-продвижения невозможно измерить одним показателем. Оценка эффективности оптимизации сайта часто имеет расплывчатые очертания. Например, даже при многократном увеличении трафика и попадании в ТОП, сложно говорить о достижении поставленных целей, если конверсия на первоначальном уровне. Поэтому перед началом работ мы выберем несколько основных показателей (KPI), на которые Вы сможете ориентироваться для оценки результата.
Создание и раскрутка сайтов, разработка и продвижение сайтов
Создание и раскрутка сайта – это комплексный процесс, состоящий из нескольких этапов. Постановка задачи, планирование и проектирование, разработка дизайна, верстка и программирование, наполнение контентом, внутренняя оптимизация, раскрутка сайта, повышение конверсии. Все этапы взаимосвязаны и только комплексный подход позволит избежать переделок и сократить сроки и трудозатраты на запуск проекта./a3478d3f49e353e.s.siteapi.org/img/25de2605b37847b31157e91a172f2603a546b3ad.PNG)
На начальном этапе крайне важно определить задачу, которую должен решить сайт и выявить ключевые показатели эффективности. Ведь управлять можно только тем, что можно измерить. Исходя из поставленных целей выбираются ключевые слова, по которым ресурс будет продвигаться. Определяются наиболее перспективные для продвижения запросы и под них готовится структура будущего сайта. Отдельные страницы и разделы сайта отводятся для раскрутки по конкретным ключевым словам. Выбираются необходимые технологии, проектируется техническое решение и определяются требования к дизайну.
Само создание сайта начинается с создания качественного и продуманного дизайна, который сегодня является обязательным условием и позволяет не только повысить конверсию посетителей в покупателей, но и улучшить пользовательские факторы сайта, влияющие на продвижение (время на сайте, количество просмотренных страниц, поведение и др.). Программирование и верстка сайта проводятся в строгом соответствии с заданием на разработку. Выполняется тестирование сайта и все выявленные недочеты устраняются. Далее наступает этап подготовки контента, наполнения и внутренней оптимизации, от качества которого зависит возможность нахождения вашего сайта в ТОП.
Выполняется тестирование сайта и все выявленные недочеты устраняются. Далее наступает этап подготовки контента, наполнения и внутренней оптимизации, от качества которого зависит возможность нахождения вашего сайта в ТОП.
Теперь ресурс готов к раскрутке в поисковых системах. Важно использовать только белые способы раскрутки, отказаться от закупки сомнительных ссылок и вести постоянную работу по повышению конверсии и качества контента. Это позволит избежать риска попадания под санкции поисковых систем и получить стабильные результаты.
Как видите, создание и раскрутка сайта является сложным процессом и важно выбрать профессиональную команду, способную эффективно решать подобные задачи.
Обращайтесь в студию 28WEB, и мы поможем создать и раскрутить ваш веб сайт или магазин! Качество и успешность ваших сайтов – это наша визитная карточка. Положительный отзыв – это наш ключевой показатель эффективности.
Дизайнеры, верстальщики, программисты и оптимизаторы работают в одной команде. Все предлагаемые нами решения прорабатываются на предмет соответствия не только техническим параметрам, но и с точки зрения влияния на раскрутку сайта.
2КомпетенцииВысокие компетенции в области управления проектами, веб дизайна, разработки и раскрутки сайтов — наше конкурентное преимущество.
3Выгодные ценыМы всегда адекватно оцениваем стоимость услуг на основе трудозатрат, необходимых на выполнение задачи. Если часть работ выполняет заказчик, то стоимость уменьшается пропорционально.
4Долговременное сотрудничество Мы осуществляем не только разработку и запуск проектов, но и продвигаем их в ТОП, оказываем услуги по поддержке и наполнению веб сайтов, консультируем клиентов.
Мы молодой, но опытный коллектив, любим свою работу и готовы браться за сайты любой сложности.
Продвижение сайтов в Москве Санкт-Петербурге и Новосибирске от 12 000 рублей
Сегодня Студия ЯЛ оказывает целый комплекс услуг по сопровождению клиентского бизнеса в сети. Мы разрабатываем сайты под ключ, занимаемся контекстной рекламой, SEO-продвижением сайтов, SMM, создаем PR-кампании, предоставляем услуги по поддержке и аналитике веб-ресурсов разных типов. Умеем работать с проектами любого масштаба, уровня сложности, тематики.
Профессионализм и опыт сотрудников Студии позволяют оперативно достигать прогнозируемых результатов и подбирать ключ к решению каждой проблемы.
Почему выбирают нас?
Наше портфолио одно из самых объемных в Рунете. К сегодняшнему дню мы реализовали более 1000 проектов по продвижению сайтов и несколько сотен – по разработке. Создавать функциональные, заточенные под получение прибыли ресурсы, выводить их по ключевым запросам в топ Яндекса и Google, настраивать контекстную рекламу, запускать эффективное SMM-продвижение, проводить грамотные аудиты – вот то, что мы умеем и любим делать.
Наверное, у вас уже назрел вопрос, кто же эти «мы»? О, у нас большая команда! В компании несколько крупных отделов, которые обычно работают над проектами сообща. Многолетняя практика доказывает, что так проще и быстрее получается выйти на нужный результат. В нашем штате есть веб-разработчики, SEO-специалисты, аналитики, контекстологи, SMM-щики, дизайнеры, копирайтеры, менеджеры. Команда опытных спецов готова удовлетворить любой запрос клиента в сфере интернет-маркетинга.
Студия ЯЛ регулярно входит в рейтинги лучших веб-студий и SEO-компаний России: Рейтинг Рунета, CMS Magazine, Ruward, «Кто продвинул?» и пр. Мы предлагаем и вам оценить результаты нашей деятельности. Загляните в раздел «Портфолио» – там вас ждет подборка интереснейших кейсов.
Для автоматизации своей работы мы создали уникальный продукт – систему Предиктор. С ее помощью удобней управлять контекстной рекламой, добиваться нужных результатов в раскрутке сайтов, отслеживать эффективность работы специалистов.
С момента основания компании мы придерживаемся собственной бизнес-модели, которую можно свести к главному правилу: «Одна ошибка – один раз». Так нам удается надежно защищать интересы своих клиентов.
Создание и продвижение сайтов — заказать в Москве, цены на разработку сайтов для СЕО в SoftMG
Создание трубо-версий основных страниц, которые загружаются быстрее – особенно актуально для мобильных устройств и мобильной выдачи Яндекса;
AMP-страницы – ускоренные страницы для мобильных устройств, разрабатываются для мобильной выдачи Гугла;
Тематические ролики о товаре, услугах и инструкции, например, по оформлению
заказа. Сейчас мало просто сфоротагрфировать товар или написать об услуге –
пользователю нужно представить информацию со всех сторон.
Сейчас мало просто сфоротагрфировать товар или написать об услуге –
пользователю нужно представить информацию со всех сторон.
Текстовые инструкции сопровождаются графическими блоками для лучшего усвоения информации. Пользователь обязательно долистает такую страницу до конца
Обеспечивает безопасный обмен данными между сайтом и компьютером пользователя, утечка информации исключена. И для Яндекса и для Гугла наличие SSL протокола – знак качества сайта.
Сервис по оптимизации скорости загрузки страниц. Наши сайты грузятся быстро – с
нами вы не будете терять пользователей, которые не дождались загрузки страницы и
ушли на другой сайт
Наши сайты грузятся быстро – с
нами вы не будете терять пользователей, которые не дождались загрузки страницы и
ушли на другой сайт
Возможность искать информацию по сайту, проговорив запрос, подобно поисковому модулю Google.
Раскрутка сайта или контекстная реклама?
Зачастую клиент задается вопросом: что выбрать раскрутку сайта по определенным целевым запросам или контекстную рекламу?
Такое противопоставление ошибочно, а подход в корне не верен. Вопрос должен состоять в том, какой метод привлечения целевого посетителя наиболее дешев и эффективен, а для этого необходим предварительный анализ целевой аудитории. Самый простой анализ можно провести по географической принадлежности и по сезонности, а также по стоимости привлечения с контекстной рекламы или раскрутки сайта.
Самый простой анализ можно провести по географической принадлежности и по сезонности, а также по стоимости привлечения с контекстной рекламы или раскрутки сайта.
По географической принадлежности – к какому региону относится наша ЦА? Получив ответ на этот вопрос, сразу же можно предположить эффективность продвижения по тем или иным запросам.
Например: Клиент – компания, оказывающая услуги дизайна интерьеров и ремонта квартир в Санкт-Петербурге. Географическая принадлежность аудитории – Петербург. Доля трафика Санкт-Петербурга примерно 10% в общей доле трафика поисковых машин. Получается что поисковое продвижение по запросам «дизайн интерьеров» и «ремонт квартир» будет на 90% бить мимо цели! Но есть ряд запросов с естественным геотаргетингом – с приставкой местности в запросе – дизайн интерьеров в Санкт-Петербурге и т.п. По ним поисковое продвижение будет на порядок менее ресурсозатратным и будет бить точно в цель – полное попадание в целевую аудиторию. Но как же быть с теми 10% петербуржцев в общей доле трафика по запросам «дизайн интерьера» и «ремонт квартир»? Здесь как раз необходима контекстная реклама по данным запросам с геотаргетингом (показом объявлений только в определенном регионе) на Петербург – если петербуржец задаст запрос «ремонт квартир» — он увидит нашу рекламу и здесь попадание рекламы в целевую аудиторию также на 100%. В случае если наша аудитория располагается в Москве и Московской области (на данный момент это порядка 55-60% трафика) – здесь стоит просчитать сколько будет стоит 1 целевой посетитель приведенный с раскрутки сайта или контекстной рекламы. Также стоит включать в данный расчет неизбежные риски поискового продвижения, поэтому если стоимость одного посетителя с контекстной рекламы и поискового продвижения будет примерно равна – стоит отдать приоритет контекстной рекламе.
Но как же быть с теми 10% петербуржцев в общей доле трафика по запросам «дизайн интерьера» и «ремонт квартир»? Здесь как раз необходима контекстная реклама по данным запросам с геотаргетингом (показом объявлений только в определенном регионе) на Петербург – если петербуржец задаст запрос «ремонт квартир» — он увидит нашу рекламу и здесь попадание рекламы в целевую аудиторию также на 100%. В случае если наша аудитория располагается в Москве и Московской области (на данный момент это порядка 55-60% трафика) – здесь стоит просчитать сколько будет стоит 1 целевой посетитель приведенный с раскрутки сайта или контекстной рекламы. Также стоит включать в данный расчет неизбежные риски поискового продвижения, поэтому если стоимость одного посетителя с контекстной рекламы и поискового продвижения будет примерно равна – стоит отдать приоритет контекстной рекламе.
По сезонности — есть ряд товаров или услуг которые имеют ярко выраженный сезонный характер, например шины или туры в Турцию. Поэтому начало поискового продвижения в разгар сезона не приведет к резкому росту продаж, т.к. максимально быстро выдвижение по самым конкурентным запросам в Яндексе составляет порядка 3 недель, а в других поисковых системах от двух месяцев. Если продвижение ресурса стартовало в начале сезона или в его разгар — стоит переориентировать бОльшие средства на контекстную рекламу, дабы получить немедленный отклик.
Поэтому начало поискового продвижения в разгар сезона не приведет к резкому росту продаж, т.к. максимально быстро выдвижение по самым конкурентным запросам в Яндексе составляет порядка 3 недель, а в других поисковых системах от двух месяцев. Если продвижение ресурса стартовало в начале сезона или в его разгар — стоит переориентировать бОльшие средства на контекстную рекламу, дабы получить немедленный отклик.
Например: Сезон продаж водонагревателей начинается в апреле и заканчивается в июле. Самый большой пик приходится на май. Если нужно немедленно увеличить продажи, а запуск кампании по продвижению сайта начался в преддверии пика, скажем, в конце апреля, то имеет смысл вложить основную массу средств в контекстную рекламу, а раскрутку сайта вести менее форсировано, но верно в верхние строчки поисковых систем, что даст приток посетителей оттуда в середине и конце сезона и поможет встретить следующий сезон в верхних строчках поисковых систем.
По стоимости привлечения одного целевого посетителя — при составлении плана продвижения вашей компании в Интернете любое рекламное агентство ОБЯЗАНО по вашей просьбе представить вам примерный план количества целевых посетителей в месяц с раскрутки сайта и их стоимость , а также по вашему желанию стоимость одного среднего целевого посетителя с контекстной рекламы. Стоит учитывать неизбежные риски, связанные с раскруткой сайта, — выпадения страниц из индекса, падение позиций и т. п. Поэтому если стоимость посетителя с контекстной рекламы будет примерно равна стоимости посетителя с поискового продвижения, стоит отдать приоритет контекстной рекламе.
Поможем с выбором и ведением рекламных каналов: проведем расчет рентабельности, составим план и покажем результат!
Например: раскрутка по группе запросов грузоперевозки для екатеринбургской компании будет гораздо дороже, нежели контекстная реклама по тем же запросам, нацеленная на ту же целевую аудиторию. Если присовокупить сюда неизбежные риски раскрутки, то становится ясно, что заказывать раскрутку сайта по данным запросам просто глупо.
Если присовокупить сюда неизбежные риски раскрутки, то становится ясно, что заказывать раскрутку сайта по данным запросам просто глупо.
Вывод: к раскрутке сайта по определенным запросам стоит прибегать только тогда, когда стоимость привлечения одного целевого посетителя будет гораздо ниже контекстной рекламы.
P.S. Раскрутка сайта всегда сопряжена с риском потери позиций и вследствие этого — трафика и продаж, поэтому всегда необходимо иметь страховочный бюджет на запуск контекстной рекламы в случае исчезновения вашего сайта с верхних строчек выдачи поисковых систем.
P.P.S. С помощью покупки контекстной рекламы можно относительно дешево охватить большое количество низкочастотных запросов, которые тяжелее охватить с помощью раскрутки, поэтому в идеале стоит всегда вести параллельно и контекстную рекламу, и раскрутку сайта по ряду запросов, дабы максимально полно охватить свою целевую аудиторию.
Полезно 2
Addgene: Promoters
Определение
Промотор — это область ДНК, в которой инициируется транскрипция гена. Промоторы являются жизненно важным компонентом векторов экспрессии, поскольку они контролируют связывание РНК-полимеразы с ДНК. РНК-полимераза транскрибирует ДНК в мРНК, которая в конечном итоге транслируется в функциональный белок. Таким образом, промоторная область контролирует, когда и где в организме экспрессируется интересующий вас ген.
Промоторы являются жизненно важным компонентом векторов экспрессии, поскольку они контролируют связывание РНК-полимеразы с ДНК. РНК-полимераза транскрибирует ДНК в мРНК, которая в конечном итоге транслируется в функциональный белок. Таким образом, промоторная область контролирует, когда и где в организме экспрессируется интересующий вас ген.
Сводка
Промоторы имеют длину около 100-1000 пар оснований и расположены рядом и обычно выше (5 ’) смысловой или кодирующей цепи транскрибируемого гена.Кодирующая цепь — это цепь ДНК, которая кодирует кодоны и последовательность которой соответствует полученному транскрипту мРНК. Антисмысловая цепь называется цепочкой-матрицей или некодирующей цепью, поскольку это цепь, которая транскрибируется РНК-полимеразой.
последовательностей ДНК, называемых элементами ответа, расположены в промоторных областях и обеспечивают стабильный сайт связывания для РНК-полимеразы и факторов транскрипции. Факторы транскрипции — это белки, которые привлекают РНК-полимеразу и контролируют и регулируют транскрипцию ДНК в мРНК.
Связывание промотора у бактерий сильно отличается от эукариот. У бактерий основной РНК-полимеразе требуется ассоциированный сигма-фактор для распознавания и связывания промотора. С другой стороны, у эукариот этот процесс намного сложнее. Эукариотам требуется минимум семь факторов транскрипции, чтобы РНК-полимераза II (эукариот-специфическая РНК-полимераза) могла связываться с промотором. Транскрипция строго контролируется как у бактерий, так и у эукариот. Промоторы контролируются различными регуляторными последовательностями ДНК, включая энхансеры, граничные элементы, инсуляторы и глушители.
Промоутеры регионов
Промотор состоит из трех основных частей: корового промотора, проксимального промотора и дистального промотора. Ниже описаны особенности этих областей в эукариотических клетках.
Основной промоутер
Центральная область промотора расположена наиболее проксимальнее стартового кодона и содержит сайт связывания РНК-полимеразы, ТАТА-бокс и сайт начала транскрипции (TSS). РНК-полимераза будет стабильно связываться с этой коровой промоторной областью, и может инициироваться транскрипция матричной цепи.ТАТА-бокс представляет собой последовательность ДНК (5′-ТАТААА-3 ‘) в центральной области промотора, где могут связываться белки общих факторов транскрипции и гистоны. Гистоны — это белки, обнаруженные в эукариотических клетках, которые упаковывают ДНК в нуклеосомы. Связывание гистонов предотвращает инициацию транскрипции, тогда как факторы транскрипции способствуют инициации транскрипции. Самая 3’-часть (ближайшая к стартовому кодону гена) основного промотора — это TSS, где фактически начинается транскрипция. Однако только эукариоты и археи содержат этот ТАТА-бокс.Большинство прокариот содержат последовательность, которая считается функционально эквивалентной, называемой боксом Прибноу, которая обычно состоит из шести нуклеотидов, TATAAT.
РНК-полимераза будет стабильно связываться с этой коровой промоторной областью, и может инициироваться транскрипция матричной цепи.ТАТА-бокс представляет собой последовательность ДНК (5′-ТАТААА-3 ‘) в центральной области промотора, где могут связываться белки общих факторов транскрипции и гистоны. Гистоны — это белки, обнаруженные в эукариотических клетках, которые упаковывают ДНК в нуклеосомы. Связывание гистонов предотвращает инициацию транскрипции, тогда как факторы транскрипции способствуют инициации транскрипции. Самая 3’-часть (ближайшая к стартовому кодону гена) основного промотора — это TSS, где фактически начинается транскрипция. Однако только эукариоты и археи содержат этот ТАТА-бокс.Большинство прокариот содержат последовательность, которая считается функционально эквивалентной, называемой боксом Прибноу, которая обычно состоит из шести нуклеотидов, TATAAT.
Проксимальный промотор
Дальше от основного промотора вы найдете проксимальный промотор, который содержит множество первичных регуляторных элементов. Проксимальный промотор находится примерно на 250 пар оснований выше TSS, и это сайт, где связываются общие факторы транскрипции.
Проксимальный промотор находится примерно на 250 пар оснований выше TSS, и это сайт, где связываются общие факторы транскрипции.
Дистальный промотор
Последняя часть области промотора называется дистальным промотором, который находится выше проксимального промотора.Дистальный промотор также содержит сайты связывания факторов транскрипции, но в основном содержит регуляторные элементы.
Эукариотические промоторы
Эукариотическая транскрипция Промоторы эукариот намного сложнее и разнообразнее, чем промоторы прокариот. Эукариотические промоторы охватывают широкий спектр последовательностей ДНК. Нет ничего необычного в наличии нескольких регуляторных элементов, таких как энхансеры, на расстоянии нескольких тысяч пар оснований от TSS. Эукариотические промоторы настолько сложны по структуре, что ДНК имеет тенденцию складываться сама по себе, что помогает объяснить, сколько физически удаленных последовательностей ДНК могут влиять на транскрипцию данного гена. ТАТА-связывающий белок связывает ТАТА-бокс и помогает в последующем связывании РНК-полимеразы. Комплекс транскрипции построен из РНК-полимеразы и нескольких белков факторов транскрипции.
ТАТА-связывающий белок связывает ТАТА-бокс и помогает в последующем связывании РНК-полимеразы. Комплекс транскрипции построен из РНК-полимеразы и нескольких белков факторов транскрипции.
Общие эукариотические промоторы, используемые в исследованиях
| Промоутер | Выражение | Описание |
|---|---|---|
| CMV | Учредительный | Сильный промотор цитомегаловируса человека млекопитающих |
| EF1a | Constituitve | Сильный промотор для млекопитающих из фактора удлинения 1 альфа человека |
| CAG | Учредительный | Сильный промотор гибридных млекопитающих |
| PGK | Учредительный | Промотор млекопитающих из гена фосфолицераткиназы |
| TRE | Индуктивный | Промотор элемента ответа на тетрациклин |
| U6 | Учредительный | Ядерный промотор U6 человека для экспрессии малой РНК |
| UAS | Особый | Промотор дрозофилы, содержащий сайты связывания Gal4 |
Бактериальные промоторы
Промоторы в бактериях содержат две короткие последовательности ДНК, расположенные в положениях -10 (10 п. н. 5 ‘или выше) и -35 от сайта начала транскрипции (TSS).Их эквивалент эукариотическому ТАТА-боксу, бокс Прибноу (ТАТААТ) расположен в позиции -10 и необходим для инициации транскрипции. Положение -35, называемое просто элементом -35, обычно состоит из последовательности TTGACA, и этот элемент контролирует скорость транскрипции. Бактериальные клетки содержат сигма-факторы, которые помогают РНК-полимеразе связываться с промоторной областью. Каждый сигма-фактор распознает разные коровые промоторные последовательности.
н. 5 ‘или выше) и -35 от сайта начала транскрипции (TSS).Их эквивалент эукариотическому ТАТА-боксу, бокс Прибноу (ТАТААТ) расположен в позиции -10 и необходим для инициации транскрипции. Положение -35, называемое просто элементом -35, обычно состоит из последовательности TTGACA, и этот элемент контролирует скорость транскрипции. Бактериальные клетки содержат сигма-факторы, которые помогают РНК-полимеразе связываться с промоторной областью. Каждый сигма-фактор распознает разные коровые промоторные последовательности.
Опероны
Хотя бактериальная транскрипция проще, чем эукариотическая транскрипция, бактерии все же имеют сложные системы регуляции генов, такие как опероны.Опероны представляют собой кластер различных генов, которые контролируются одним промотором и оператором. Опероны распространены у прокайотов, особенно у бактерий, но также были обнаружены у эукариот. Опероны состоят из промотора, который распознается РНК-полимеразой, оператора, сегмента ДНК, с которым может связываться репрессор или активатор, и структурных генов, которые транскрибируются вместе.
Регуляция оперона может быть как отрицательной, так и положительной. Отрицательные репрессивные опероны обычно связываются репрессорным белком, который предотвращает транскрипцию.Когда молекула-индуктор связывается с репрессором, она меняет свою конформацию, предотвращая связывание с оператором и тем самым обеспечивая возможность транскрипции. Оперон Lac у бактерий является примером отрицательно контролируемого оперона.
Положительный репрессируемый оперон работает противоположным образом. Оперон обычно транскрибируется до тех пор, пока репрессор / корепрессор не свяжется с оператором, предотвращая транскрипцию. Оперон trp, участвующий в производстве триптофана, является примером положительно контролируемого оперона.
Общие бактериальные промоторы, использованные в исследованиях
| Промоутер | Выражение | Описание |
|---|---|---|
| T7 | Конститутивная, но требует РНК-полимеразы Т7 | Промотор бактериофага Т7 |
| Сп6 | Конститутивная, но требует РНК-полимеразы Sp6 | Промотор бактериофага Sp6 |
| лак | Конститутивное при отсутствии lac-репрессора (lacI или lacIq). Может индуцироваться IPTG или лактозой Может индуцироваться IPTG или лактозой | Промоутер от Lac operon |
| араБад | Индуцируется арабинозой | Промотор метаболического оперона арабинозы |
| т.р. | Подавляемость триптофаном | Промотор из E. coli триптофановый оперон |
| Ptac | Регулируется как промотор лака | Гибридный промотор lac и trp |
Типы РНК-полимераз
Промоторы контролируют связывание РНК-полимеразы с ДНК, чтобы инициировать транскрипцию генов.Существует три типа РНК-полимераз, которые транскрибируют разные гены.
РНК-полимераза I транскрибирует гены, кодирующие рибосомную РНК (рРНК), которая является основным компонентом структуры рибосомы клетки. Рибосомы — это место синтеза белка, где мРНК транслируется в белок.
Рибосомы — это место синтеза белка, где мРНК транслируется в белок.
РНК-полимераза II транскрибирует информационную РНК (мРНК), которая является РНК, ответственной за обеспечение стабильной матрицы для трансляции белка.
РНК-полимераза III транскрибирует гены, кодирующие транспортную РНК (тРНК), адапторные молекулы, которые отвечают за перенос аминокислот в рибосому при синтезе белков. РНК-полимераза III также транскрибирует малые РНК, такие как shRNA и gRNA.
Ресурсы
БлогAddgene, включая нашу популярную серию Plasmids 101, охватывает самые разные темы — от новейших достижений в плазмидных технологиях и исследованиях до обзоров основ молекулярной биологии и компонентов плазмид.
Промоутерский регион у эукариот и прокариот
Разница между эукариотами и Прокариотические промоторы
Прокариотические промоторы
У прокариот промотор состоит из двух
короткие последовательности в положениях -10 и -35 перед
сайт начала транскрипции.
- Последовательность -10 называется Pribnow box, или элемент -10, и обычно состоит из шесть нуклеотидов TATAAT. Ящик Прибнов абсолютно необходимо для начала транскрипции прокариот.
- Другая последовательность — 35 (-35 element) обычно состоит из шести нуклеотидов TTGACA. Его присутствие обеспечивает очень высокую скорость транскрипции.
Промоторы эукариот
Эукариотические промоторы чрезвычайно разнообразны
и их трудно охарактеризовать.Обычно они лежат выше по течению
гена и может иметь регуляторные элементы в несколько тысяч оснований
вдали от сайта начала транскрипции. У эукариот
транскрипционный комплекс может привести к изгибу ДНК
сам по себе, что позволяет размещать регуляторные последовательности далеко
с фактического сайта транскрипции. Многие эукариотические
промоторы, содержат ТАТА-бокс (последовательность TATAAA ), который, в свою очередь, связывает привязку TATA
белок, который помогает в образовании РНК-полимеразы
транскрипционный комплекс.Коробка TATA обычно лежит очень
близко к сайту начала транскрипции (часто в пределах 50
баз).
Многие эукариотические
промоторы, содержат ТАТА-бокс (последовательность TATAAA ), который, в свою очередь, связывает привязку TATA
белок, который помогает в образовании РНК-полимеразы
транскрипционный комплекс.Коробка TATA обычно лежит очень
близко к сайту начала транскрипции (часто в пределах 50
баз).
Регулирование эукариотических генов | Безграничная биология
Промоутер и машина для транскрипции
Когда факторы транскрипции связываются с промоторной областью, РНК-полимераза размещается в ориентации, позволяющей начать транскрипцию.
Цели обучения
Опишите роль промоторов в транскрипции РНК
Основные выводы
Ключевые моменты
- Промотор предназначен для связывания факторов транскрипции, которые контролируют инициацию транскрипции.
- Промоторная область может быть короткой или довольно длинной; чем длиннее промотор, тем больше свободного пространства для связывания белков.
- Чтобы инициировать транскрипцию, фактор транскрипции (TFIID) связывается с боксом ТАТА, что заставляет другие факторы транскрипции впоследствии связываться с боксом ТАТА.
- После сборки комплекса инициации транскрипции РНК-полимераза может связываться со своей вышестоящей последовательностью и затем фосфорилироваться.
- Фосфорилирование РНК-полимеразы высвобождает часть белка из ДНК для активации комплекса инициации транскрипции и помещает РНК-полимеразу в правильную ориентацию для начала транскрипции.
- Факторы транскрипции реагируют на стимулы окружающей среды, которые заставляют белки находить свои сайты связывания и инициировать транскрипцию необходимого гена.
Ключевые термины
- ТАТА-бокс : последовательность ДНК (цис-регуляторный элемент), обнаруженная в промоторной области генов у архей и эукариот
- фактор транскрипции : белок, который связывается со специфическими последовательностями ДНК, тем самым контролируя поток (или транскрипцию) генетической информации от ДНК к мРНК
- промотор : участок ДНК, который контролирует инициацию транскрипции РНК
Промоутер и устройство для транскрипции
Гены организованы таким образом, чтобы облегчить контроль экспрессии генов. Промоторная область находится непосредственно перед кодирующей последовательностью. Этот участок может быть коротким (всего несколько нуклеотидов в длину) или довольно длинным (длиной в сотни нуклеотидов). Чем длиннее промотор, тем больше свободного пространства для связывания белков. Это также добавляет больше контроля над процессом транскрипции. Длина промотора зависит от гена и может сильно различаться между генами. Следовательно, уровень контроля экспрессии генов также может сильно различаться между генами. Промотор предназначен для связывания факторов транскрипции, которые контролируют инициацию транскрипции.
Промоторная область находится непосредственно перед кодирующей последовательностью. Этот участок может быть коротким (всего несколько нуклеотидов в длину) или довольно длинным (длиной в сотни нуклеотидов). Чем длиннее промотор, тем больше свободного пространства для связывания белков. Это также добавляет больше контроля над процессом транскрипции. Длина промотора зависит от гена и может сильно различаться между генами. Следовательно, уровень контроля экспрессии генов также может сильно различаться между генами. Промотор предназначен для связывания факторов транскрипции, которые контролируют инициацию транскрипции.
В промоторной области, прямо перед сайтом старта транскрипции, находится ТАТА-бокс. Этот блок представляет собой просто повторение динуклеотидов тимина и аденина (буквально, повторы ТАТА). РНК-полимераза связывается с комплексом инициации транскрипции, позволяя транскрипции происходить. Чтобы инициировать транскрипцию, фактор транскрипции (TFIID) первым связывается с боксом TATA. Связывание TFIID задействует другие факторы транскрипции, включая TFIIB, TFIIE, TFIIF и TFIIH, в бокс TATA.Как только этот комплекс инициации транскрипции собран, РНК-полимераза может связываться со своей вышестоящей последовательностью. При связывании с факторами транскрипции РНК-полимераза фосфорилируется. Это высвобождает часть белка из ДНК, чтобы активировать комплекс инициации транскрипции, и помещает РНК-полимеразу в правильную ориентацию для начала транскрипции; Изгибающий ДНК белок приводит энхансер, который может находиться на значительном расстоянии от гена, в контакт с факторами транскрипции и белками-медиаторами.
Связывание TFIID задействует другие факторы транскрипции, включая TFIIB, TFIIE, TFIIF и TFIIH, в бокс TATA.Как только этот комплекс инициации транскрипции собран, РНК-полимераза может связываться со своей вышестоящей последовательностью. При связывании с факторами транскрипции РНК-полимераза фосфорилируется. Это высвобождает часть белка из ДНК, чтобы активировать комплекс инициации транскрипции, и помещает РНК-полимеразу в правильную ориентацию для начала транскрипции; Изгибающий ДНК белок приводит энхансер, который может находиться на значительном расстоянии от гена, в контакт с факторами транскрипции и белками-медиаторами.
Промоторы : показан обобщенный промотор гена, транскрибируемого РНК-полимеразой II. Факторы транскрипции распознают промотор. Затем РНК-полимераза II связывается и образует комплекс инициации транскрипции.
Помимо общих факторов транскрипции, с промотором могут связываться другие факторы транскрипции, регулирующие транскрипцию гена. Эти факторы транскрипции связываются с промоторами определенного набора генов. Они не являются общими факторами транскрипции, которые связываются с каждым комплексом промотора, но задействованы в определенной последовательности на промоторе определенного гена.В клетке есть сотни факторов транскрипции, каждый из которых специфически связывается с определенным мотивом последовательности ДНК. Когда факторы транскрипции связываются с промотором непосредственно перед кодируемым геном, их называют цис-действующими элементами, потому что они находятся на одной хромосоме, рядом с геном. Область, с которой связывается конкретный фактор транскрипции, называется сайтом связывания фактора транскрипции. Факторы транскрипции реагируют на стимулы окружающей среды, которые заставляют белки находить свои сайты связывания и инициировать транскрипцию необходимого гена.
Эти факторы транскрипции связываются с промоторами определенного набора генов. Они не являются общими факторами транскрипции, которые связываются с каждым комплексом промотора, но задействованы в определенной последовательности на промоторе определенного гена.В клетке есть сотни факторов транскрипции, каждый из которых специфически связывается с определенным мотивом последовательности ДНК. Когда факторы транскрипции связываются с промотором непосредственно перед кодируемым геном, их называют цис-действующими элементами, потому что они находятся на одной хромосоме, рядом с геном. Область, с которой связывается конкретный фактор транскрипции, называется сайтом связывания фактора транскрипции. Факторы транскрипции реагируют на стимулы окружающей среды, которые заставляют белки находить свои сайты связывания и инициировать транскрипцию необходимого гена.
Усилители и репрессоры транскрипции
Энхансеры увеличивают скорость транскрипции генов, а репрессоры снижают скорость транскрипции.
Цели обучения
Объясните, как энхансеры и репрессоры регулируют экспрессию генов
Основные выводы
Ключевые моменты
- Энхансеры могут быть расположены выше гена, в кодирующей области гена, ниже гена или за тысячи нуклеотидов.
- Когда белок, изгибающий ДНК, связывается с энхансером, форма ДНК изменяется, что позволяет взаимодействовать между активаторами и факторами транскрипции.
- Репрессоры реагируют на внешние стимулы, предотвращая связывание активирующих факторов транскрипции.
- Корепрессоры могут подавлять инициацию транскрипции, рекрутируя гистондеацетилазу.
- Деактилирование гистонов увеличивает положительный заряд на гистонах, что усиливает взаимодействие между гистонами и ДНК, делая ДНК менее доступной для транскрипции.
Ключевые термины
- энхансер : короткая область ДНК, которая может увеличивать транскрипцию генов
- репрессор : любой белок, который связывается с ДНК и, таким образом, регулирует экспрессию генов, снижая скорость транскрипции
- активатор : любое химическое вещество или агент, который регулирует один или несколько генов путем увеличения скорости транскрипции
Энхансеры и транскрипция
В некоторых эукариотических генах есть участки, которые помогают увеличить или усилить транскрипцию. Эти области, называемые энхансерами, не обязательно близки к генам, которые они усиливают. Они могут быть расположены выше гена, в кодирующей области гена, ниже гена или могут находиться на расстоянии тысяч нуклеотидов.
Эти области, называемые энхансерами, не обязательно близки к генам, которые они усиливают. Они могут быть расположены выше гена, в кодирующей области гена, ниже гена или могут находиться на расстоянии тысяч нуклеотидов.
Энхансерные области представляют собой связывающие последовательности или сайты для факторов транскрипции. Когда белок, изгибающий ДНК, связывается с энхансером, форма ДНК изменяется. Это изменение формы позволяет взаимодействовать между активаторами, связанными с энхансерами, и факторами транскрипции, связанными с промоторной областью, и РНК-полимеразой.В то время как ДНК обычно изображается как прямая линия в двух измерениях, на самом деле это трехмерный объект. Следовательно, нуклеотидная последовательность на тысячи нуклеотидов может сворачиваться и взаимодействовать со специфическим промотором.
Энхансеры : Энхансер — это последовательность ДНК, которая способствует транскрипции. Каждый энхансер состоит из коротких последовательностей ДНК, называемых дистальными элементами управления. Активаторы, связанные с дистальными элементами управления, взаимодействуют с белками-медиаторами и факторами транскрипции.
Выключение генов: репрессоры транскрипции
Подобно прокариотическим клеткам, эукариотические клетки также обладают механизмами предотвращения транскрипции. Репрессоры транскрипции могут связываться с областями промотора или энхансера и блокировать транскрипцию. Подобно активаторам транскрипции, репрессоры реагируют на внешние стимулы, предотвращая связывание активирующих факторов транскрипции.
Корепрессор — это белок, который снижает экспрессию гена за счет связывания с фактором транскрипции, который содержит ДНК-связывающий домен.Корепрессор сам по себе не может связывать ДНК. Корепрессор может подавлять инициацию транскрипции путем привлечения гистондеацетилазы, которая катализирует удаление ацетильных групп с остатков лизина. Это увеличивает положительный заряд на гистонах, что усиливает взаимодействие между гистонами и ДНК, делая ДНК менее доступной для процесса транскрипции.
Эпигенетический контроль: регулирование доступа к генам в хромосоме
Как упаковка ДНК вокруг гистоновых белков, так и химические модификации ДНК или белков могут изменять экспрессию генов.
Цели обучения
Обсудите, как регуляция эукариотических генов происходит на эпигенетическом уровне и различные эпигенетические изменения, которые могут быть внесены в ДНК.
Основные выводы
Ключевые моменты
- ДНК упаковывается путем обертывания гистоновых белков в структуры, называемые нуклеосомами, которые напоминают бусинки на нитке.
- Когда ДНК должна быть расшифрована, нуклеосомы могут ускользнуть от этой области ДНК, открывая ее для транскрипционного аппарата клетки.
- Химические модификации гистоновых белков или самой ДНК сигнализируют, должна ли конкретная область генома быть «открытой» или «закрытой» для транскрипционного аппарата.
- Модификации, такие как ацетилирование или метилирование гистонов, могут изменить то, насколько плотно ДНК обернута вокруг них, в то время как метилирование ДНК изменяет то, как ДНК взаимодействует с белками, включая гистоновые белки, которые контролируют доступ к области.
- Этот тип генетической регуляции называется эпигенетической регуляцией («выше генетики»), поскольку он не изменяет нуклеотидную последовательность ДНК.

Ключевые термины
- нуклеосома : любая из субъединиц, которая повторяется в хроматине; спираль ДНК, окружающая гистоновое ядро
- эпигенетика : исследование наследственных изменений, вызванных активацией и деактивацией генов без каких-либо изменений в последовательности ДНК
- гистон : любой из различных простых водорастворимых белков, которые богаты основными аминокислотами лизином и аргинином и образуют комплекс с ДНК в нуклеосомах эукариотического хроматина
Эпигенетический контроль: регулирование доступа к генам в хромосоме
Геном человека кодирует более 20 000 генов; каждая из 23 пар хромосом человека кодирует тысячи генов.ДНК в ядре точно наматывается, складывается и уплотняется в хромосомы, так что она помещается в ядро. Он также организован таким образом, что определенные сегменты могут быть доступны по мере необходимости для определенного типа ячеек.
Он также организован таким образом, что определенные сегменты могут быть доступны по мере необходимости для определенного типа ячеек.
Первый уровень организации или упаковки — это наматывание цепей ДНК вокруг гистоновых белков. Гистоны упаковывают и упорядочивают ДНК в структурные единицы, называемые нуклеосомными комплексами, которые могут контролировать доступ белков к участкам ДНК. Под электронным микроскопом эта намотка ДНК вокруг гистоновых белков с образованием нуклеосом выглядит как маленькие бусинки на нитке.Эти шарики (гистоновые белки) могут перемещаться по нити (ДНК) и изменять структуру молекулы.
Упаковка ДНК : ДНК складывается вокруг гистоновых белков для создания (а) нуклеосомных комплексов. Эти нуклеосомы контролируют доступ белков к лежащей в основе ДНК. При просмотре в электронный микроскоп (б) нуклеосомы выглядят как бусинки на нитке.
Если ДНК, кодирующая определенный ген, должна быть транскрибирована в РНК, нуклеосомы, окружающие эту область ДНК, могут скользить вниз по ДНК, чтобы открыть эту специфическую хромосомную область и позволить транскрипционному аппарату (РНК-полимеразе) инициировать транскрипцию. Нуклеосомы могут двигаться, открывая структуру хромосомы, чтобы обнажить сегмент ДНК, но делать это очень контролируемым образом.
Нуклеосомы могут двигаться, открывая структуру хромосомы, чтобы обнажить сегмент ДНК, но делать это очень контролируемым образом.
Нуклеосомы могут менять положение, чтобы обеспечить транскрипцию генов. : Нуклеосомы могут скользить по ДНК. Когда нуклеосомы расположены близко друг к другу (вверху), факторы транскрипции не могут связываться, и экспрессия генов выключена. Когда нуклеосомы расположены далеко друг от друга (внизу), обнажается ДНК. Факторы транскрипции могут связываться, обеспечивая экспрессию генов.Модификации гистонов и ДНК влияют на расстояние между нуклеосомами.
Как движутся гистоновые белки, зависит от сигналов, обнаруживаемых как гистоновыми белками, так и ДНК. Эти сигналы представляют собой метки или модификации, добавленные к гистоновым белкам и ДНК, которые сообщают гистонам, должна ли хромосомная область быть открытой или закрытой. Эти теги не являются постоянными, но могут быть добавлены или удалены по мере необходимости. Это химические модификации (фосфатные, метильные или ацетильные группы), которые присоединяются к определенным аминокислотам в белке или к нуклеотидам ДНК. Метки не изменяют последовательность оснований ДНК, но они изменяют, насколько плотно ДНК намотана вокруг гистоновых белков. ДНК — это отрицательно заряженная молекула; следовательно, изменения заряда гистона изменят то, насколько плотно будет намотана молекула ДНК. В неизмененном виде гистоновые белки имеют большой положительный заряд; за счет добавления химических модификаций, таких как ацетильные группы, заряд становится менее положительным.
Метки не изменяют последовательность оснований ДНК, но они изменяют, насколько плотно ДНК намотана вокруг гистоновых белков. ДНК — это отрицательно заряженная молекула; следовательно, изменения заряда гистона изменят то, насколько плотно будет намотана молекула ДНК. В неизмененном виде гистоновые белки имеют большой положительный заряд; за счет добавления химических модификаций, таких как ацетильные группы, заряд становится менее положительным.
Модификации гистонов и ДНК могут изменять экспрессию генов : гистоновые белки и нуклеотиды ДНК можно модифицировать химически.Модификации влияют на расстояние между нуклеосомами и экспрессию генов.
Сама молекула ДНК также может быть модифицирована. Это происходит в очень специфических областях, называемых островками CpG. Это участки с высокой частотой пар цитозиновых и гуаниндинуклеотидных ДНК (CG), обнаруженных в промоторных областях генов. Когда такая конфигурация существует, цитозиновый член пары может быть метилирован (добавлена метильная группа). Эта модификация изменяет способ взаимодействия ДНК с белками, включая гистоновые белки, которые контролируют доступ к этой области.Сильно метилированные (гиперметилированные) участки ДНК с деацетилированными гистонами плотно скручены и транскрипционно неактивны. Эти изменения в ДНК передаются от родителей к потомкам, так что, хотя последовательность ДНК не изменяется, образец экспрессии генов передается следующему поколению.
Эта модификация изменяет способ взаимодействия ДНК с белками, включая гистоновые белки, которые контролируют доступ к этой области.Сильно метилированные (гиперметилированные) участки ДНК с деацетилированными гистонами плотно скручены и транскрипционно неактивны. Эти изменения в ДНК передаются от родителей к потомкам, так что, хотя последовательность ДНК не изменяется, образец экспрессии генов передается следующему поколению.
Этот тип регуляции генов называется эпигенетической регуляцией. Эпигенетика означает «выше генетики». Изменения, происходящие с белками гистонов и ДНК, не изменяют нуклеотидную последовательность и не являются постоянными.Напротив, эти изменения являются временными (хотя они часто сохраняются в течение нескольких раундов клеточного деления) и изменяют хромосомную структуру (открытая или закрытая) по мере необходимости. Ген может быть включен или выключен в зависимости от местоположения и модификаций гистоновых белков и ДНК. Если ген должен быть транскрибирован, гистоновые белки и ДНК модифицируются вокруг хромосомной области, кодирующей этот ген. Это открывает хромосомную область для доступа РНК-полимеразы и других белков, называемых факторами транскрипции, для связывания с промоторной областью, расположенной прямо перед геном, и инициирования транскрипции.Если ген должен оставаться выключенным или заглушенным, гистоновые белки и ДНК имеют разные модификации, которые сигнализируют о закрытой хромосомной конфигурации. В этой закрытой конфигурации РНК-полимераза и факторы транскрипции не имеют доступа к ДНК, и транскрипция не может происходить.
Это открывает хромосомную область для доступа РНК-полимеразы и других белков, называемых факторами транскрипции, для связывания с промоторной областью, расположенной прямо перед геном, и инициирования транскрипции.Если ген должен оставаться выключенным или заглушенным, гистоновые белки и ДНК имеют разные модификации, которые сигнализируют о закрытой хромосомной конфигурации. В этой закрытой конфигурации РНК-полимераза и факторы транскрипции не имеют доступа к ДНК, и транскрипция не может происходить.
Сплайсинг РНК
Сплайсинг РНКпозволяет производить множество изоформ белка из одного гена путем удаления интронов и объединения разных экзонов.
Цели обучения
Объясните роль сплайсинга РНК в регуляции экспрессии генов
Основные выводы
Ключевые моменты
- Интроны — это промежуточные последовательности в молекуле пре-мРНК, которые не кодируют белки и удаляются во время процессинга РНК сплайсосомой.
- Экзоны представляют собой экспрессирующие последовательности в молекуле пре-мРНК, которые сращиваются вместе после удаления интронов с образованием зрелых молекул мРНК, которые транслируются в белки.
- Альтернативный сплайсинг позволяет производить различные изоформы белка из одного кодирующего гена.
- Сплайсосома — это комплекс, состоящий как из молекул РНК, так и из белков, которые определяют, какие интроны исключить, а какие экзоны сохранить и связать вместе.

Ключевые термины
- интрон : часть расщепленного гена, которая включается в транскрипты пре-РНК, но удаляется во время процессинга РНК и быстро разрушается
- экзон : область транскрибируемого гена, присутствующего в конечной функциональной молекуле РНК
- сплайсосома : динамический комплекс РНК и белковых субъединиц, который удаляет интроны из предшественника мРНК
Сплайсинг РНК, первый этап посттранскрипционного контроля
Экспрессия гена — это процесс передачи генетической информации от гена, состоящего из ДНК, в функциональный генный продукт, состоящий из РНК или белка.Генетическая информация передается от ДНК к РНК в процессе транскрипции, а затем от РНК к белку в процессе трансляции. Чтобы гарантировать получение нужных продуктов, экспрессия генов регулируется на многих различных этапах во время транскрипции и трансляции и между ними. У эукариот ген содержит дополнительные последовательности, которые не кодируют белок. У этих организмов транскрипция ДНК производит пре-мРНК. Эти пре-мРНК-транскрипты часто содержат области, называемые интронами, которые представляют собой промежуточные последовательности, которые необходимо удалить перед трансляцией в процессе сплайсинга.Области РНК, кодирующие белок, называются экзонами. Сплайсинг можно регулировать таким образом, чтобы разные мРНК могли содержать или не содержать экзоны, в процессе, называемом альтернативным сплайсингом. Альтернативный сплайсинг позволяет продуцировать более одного белка из гена и является важным регуляторным этапом при определении того, какие функциональные белки образуются в результате экспрессии гена. Таким образом, сплайсинг — это первая стадия посттранскрипционного контроля.
Чтобы гарантировать получение нужных продуктов, экспрессия генов регулируется на многих различных этапах во время транскрипции и трансляции и между ними. У эукариот ген содержит дополнительные последовательности, которые не кодируют белок. У этих организмов транскрипция ДНК производит пре-мРНК. Эти пре-мРНК-транскрипты часто содержат области, называемые интронами, которые представляют собой промежуточные последовательности, которые необходимо удалить перед трансляцией в процессе сплайсинга.Области РНК, кодирующие белок, называются экзонами. Сплайсинг можно регулировать таким образом, чтобы разные мРНК могли содержать или не содержать экзоны, в процессе, называемом альтернативным сплайсингом. Альтернативный сплайсинг позволяет продуцировать более одного белка из гена и является важным регуляторным этапом при определении того, какие функциональные белки образуются в результате экспрессии гена. Таким образом, сплайсинг — это первая стадия посттранскрипционного контроля.
Альтернативное сращивание : Существует пять основных режимов альтернативного сращивания.
Альтернативный сплайсинг : Пре-мРНК можно альтернативно сплайсировать для создания различных белков.
Альтернативная сварка
Альтернативный сплайсинг — это процесс, который происходит во время экспрессии гена и позволяет производить множество белков (изоформ белков) из одного кодирующего гена. Альтернативный сплайсинг может происходить из-за различных способов, которыми экзон может быть исключен из информационной РНК или включен в нее. Это также может произойти, если части экзона исключены / включены или если есть включение интронов.Например, если пре-мРНК имеет четыре экзона (A, B, C и D), их можно сплайсировать и транслировать в различных комбинациях. Экзоны A, B и C можно транслировать вместе, или экзоны A, C и D можно транслировать. В результате получается так называемое альтернативное сращивание. Характер сплайсинга и продуцирования альтернативно сплайсированной информационной РНК контролируется связыванием регуляторных белков (транс-действующих белков, содержащих гены) с цис-действующими сайтами, которые находятся на пре-РНК. Некоторые из этих регуляторных белков включают активаторы сплайсинга (белки, которые способствуют определенным сайтам сплайсинга) и репрессоры сплайсинга (белки, которые уменьшают использование определенных сайтов). Некоторые распространенные репрессоры сплайсинга включают: гетерогенный ядерный рибонуклеопротеин (hnRNP) и белок, связывающий полипиримидиновый тракт (PTB). Белки, которые транслируются из матричных РНК, подвергнутых альтернативному сплайсингу, различаются последовательностью аминокислот, что приводит к изменению функции белка. Это одна из причин, по которой геном человека может кодировать большое разнообразие белков.Альтернативный сплайсинг — распространенный процесс, который происходит у эукариот; большинство мультиэкзонных генов у человека сплайсируются альтернативно. К сожалению, аномальные вариации сплайсинга также являются причиной многих генетических заболеваний и нарушений.
Некоторые из этих регуляторных белков включают активаторы сплайсинга (белки, которые способствуют определенным сайтам сплайсинга) и репрессоры сплайсинга (белки, которые уменьшают использование определенных сайтов). Некоторые распространенные репрессоры сплайсинга включают: гетерогенный ядерный рибонуклеопротеин (hnRNP) и белок, связывающий полипиримидиновый тракт (PTB). Белки, которые транслируются из матричных РНК, подвергнутых альтернативному сплайсингу, различаются последовательностью аминокислот, что приводит к изменению функции белка. Это одна из причин, по которой геном человека может кодировать большое разнообразие белков.Альтернативный сплайсинг — распространенный процесс, который происходит у эукариот; большинство мультиэкзонных генов у человека сплайсируются альтернативно. К сожалению, аномальные вариации сплайсинга также являются причиной многих генетических заболеваний и нарушений.
Механизм сплайсинга : Альтернативный сплайсинг может приводить к образованию изоформ белка.
Сплайсосома
Сплайсинг информационной РНК осуществляется и катализируется макромолекулярным комплексом, известным как сплайсосома.Области лигирования и расщепления определяются множеством субъединиц сплайсосомы, которые включают сайт разветвления (A) и сайты 5 ‘и 3’ сплайсинга. Взаимодействия между этими субъединицами и малыми ядерными рибонуклеопротеинами (snRNP), обнаруженными в сплайсосоме, создают комплекс сплайсосомы A, который помогает определить, какие интроны исключить, а какие экзоны сохранить и связать вместе. После того, как интроны расщеплены и удалены, экзоны соединяются фосфодиэфирной связью.
Регуляторные белки
Как отмечалось выше, сплайсинг регулируется белками-репрессорами и белками-активаторами, которые также известны как транс-действующие белки.Не менее важны сайленсеры и энхансеры, которые обнаруживаются на информационных РНК, также известных как цис-действующие сайты. Эти регулирующие функции работают вместе, чтобы создать код сращивания, который определяет альтернативное сращивание.
Комплекс инициации и скорость трансляции
Первым этапом трансляции является сборка рибосомы, для которой требуются факторы инициации.
Цели обучения
Обсудить, как эукариоты собирают рибосомы на мРНК, чтобы начать трансляцию
Основные выводы
Ключевые моменты
- Компоненты, участвующие в сборке рибосом, объединяются с помощью белков, называемых факторами инициации, которые связываются с небольшой субъединицей рибосомы.
- Инициаторная тРНК используется для определения местоположения стартового кодона AUG (аминокислота метионин), который устанавливает рамку считывания для цепи мРНК.
- GTP, переносимый eIF2, является источником энергии, используемым для загрузки инициаторной тРНК, переносимой малой субъединицей рибосомы на правильном стартовом кодоне в мРНК.
- GTP, переносимый eIF5, является источником энергии для сборки вместе больших и малых рибосомных субъединиц.
Ключевые термины
- рамка считывания : любой из трех возможных триплетов кодонов, в которых может быть транскрибирована последовательность ДНК
- фосфорилирование : добавление фосфатной группы к соединению; часто катализируется ферментами
Скорость сборки и трансляции рибосом
Подобно транскрипции, трансляция контролируется белками, которые связываются и запускают процесс. В переводе, прежде чем синтез белка может начаться, сборка рибосомы должна быть завершена. Это многоэтапный процесс.
В переводе, прежде чем синтез белка может начаться, сборка рибосомы должна быть завершена. Это многоэтапный процесс.
При сборке рибосомы большая и малая субъединицы рибосомы и инициаторная тРНК (тРНК и ), содержащая первую аминокислоту конечной полипептидной цепи, все объединяются в кодоне начала трансляции на мРНК, чтобы позволить начать трансляцию. Во-первых, малая субъединица рибосомы связывается с тРНК –, которая несет метионин у эукариот и архей и несет N-формилметионин в бактериях.(Поскольку тРНК i несет аминокислоту, она считается заряженной.) Затем небольшая рибосомная субъединица с заряженной тРНК i все еще связанная сканирует вдоль цепи мРНК, пока не достигнет стартового кодона AUG, который указывает, где начнется перевод. Стартовый кодон также устанавливает рамку считывания для цепи мРНК, которая имеет решающее значение для синтеза правильной последовательности аминокислот. Сдвиг рамки считывания приводит к неправильной трансляции мРНК. Затем антикодон на тРНК и связывается со стартовым кодоном посредством спаривания оснований.Комплекс, состоящий из мРНК, заряженной тРНК i и малой субъединицы рибосомы, прикрепляется к большой субъединице рибосомы, что завершает сборку рибосомы. Эти компоненты объединяются с помощью белков, называемых факторами инициации, которые связываются с небольшой рибосомной субъединицей во время инициации и обнаруживаются во всех трех сферах жизни. Кроме того, клетка тратит энергию GTP, чтобы помочь сформировать комплекс инициации. Как только сборка рибосомы завершена, заряженная тРНК i позиционируется в Р-сайте рибосомы, а пустой А-сайт готов для следующей аминоацил-тРНК.Синтез полипептида начинается и всегда продолжается от N-конца к C-концу, что называется направлением от N к C.
Затем антикодон на тРНК и связывается со стартовым кодоном посредством спаривания оснований.Комплекс, состоящий из мРНК, заряженной тРНК i и малой субъединицы рибосомы, прикрепляется к большой субъединице рибосомы, что завершает сборку рибосомы. Эти компоненты объединяются с помощью белков, называемых факторами инициации, которые связываются с небольшой рибосомной субъединицей во время инициации и обнаруживаются во всех трех сферах жизни. Кроме того, клетка тратит энергию GTP, чтобы помочь сформировать комплекс инициации. Как только сборка рибосомы завершена, заряженная тРНК i позиционируется в Р-сайте рибосомы, а пустой А-сайт готов для следующей аминоацил-тРНК.Синтез полипептида начинается и всегда продолжается от N-конца к C-концу, что называется направлением от N к C.
У эукариот несколько белков эукариотических факторов инициации (eIFs) участвуют в сборке рибосом. Фактор инициации эукариот-2 (eIF-2) активен, когда он связывается с гуанозинтрифосфатом (GTP). Со связанным с ним GTP, белок eIF-2 связывается с небольшой 40S субъединицей рибосомы. Затем тРНК инициатора, заряженная метионином (Met-tRNA i ), связывается с рибосомным комплексом GTP-eIF-2 / 40S, и как только все эти компоненты связываются друг с другом, их вместе называют комплексом 43S.
Со связанным с ним GTP, белок eIF-2 связывается с небольшой 40S субъединицей рибосомы. Затем тРНК инициатора, заряженная метионином (Met-tRNA i ), связывается с рибосомным комплексом GTP-eIF-2 / 40S, и как только все эти компоненты связываются друг с другом, их вместе называют комплексом 43S.
Факторы инициации эукариот eIF1, eIF3, eIF4 и eIF5 помогают транслировать комплекс 43S в 5′-m 7 G cap мРНК. После связывания с кэпом 5’m 7 G мРНК комплекс 43S начинает двигаться вниз по мРНК, пока не достигнет инициирующего кодона AUG в начале рамки считывания мРНК. Последовательности вокруг AUG могут помочь гарантировать, что правильный AUG используется в качестве инициирующего кодона в мРНК.
Как только комплекс 43S находится в инициирующем AUG, tRNAi-Met располагается над AUG.Антикодон на тРНК i -Met пары оснований с кодоном AUG. В этот момент GTP, связанный с eIF2 в комплексе 43S, гидролизуется до GDP + фосфата, и высвобождается энергия. Эта энергия используется для высвобождения eIF2 (со связанным с ним GDP) из комплекса 43S, оставляя рибосомную субъединицу 40S и тРНК i -Met в сайте начала трансляции мРНК.
Эта энергия используется для высвобождения eIF2 (со связанным с ним GDP) из комплекса 43S, оставляя рибосомную субъединицу 40S и тРНК i -Met в сайте начала трансляции мРНК.
Далее, eIF5 со связанным GTP связывается с субъединицей рибосомы 40S, входящей в комплекс с мРНК и тРНК i -Met.EIF5-GTP позволяет связываться большой рибосомной субъединице 60S. Как только субъединица рибосомы 60S прибывает, eIF5 гидролизует связанный с ней GTP до GDP + фосфата, и высвобождается энергия. Эта энергия обеспечивает сборку двух рибосомных субъединиц в интактную 80S рибосому с tRNAi-Met в ее P-сайте, а также в сочетании с инициирующим кодоном AUG на мРНК. Перевод готов начать.
Связывание eIF-2 с субъединицей рибосомы 40S контролируется фосфорилированием. Если eIF-2 фосфорилируется, он претерпевает конформационные изменения и не может связываться с GTP.Следовательно, комплекс 43S не может сформироваться должным образом, и трансляция затруднена. Когда eIF-2 остается нефосфорилированным, он связывает 40S субъединицу рибосомы и активно транслирует белок.
Комплекс инициации трансляции : Экспрессия гена может контролироваться факторами, которые связывают комплекс инициации трансляции.
Способность полностью собрать рибосому напрямую влияет на скорость, с которой происходит трансляция. Но синтез белка также регулируется на различных других уровнях, включая синтез мРНК, синтез тРНК, синтез рРНК и синтез эукариотических факторов инициации.Изменение любого из этих компонентов влияет на скорость, с которой может происходить перевод.
Регулирование активности и долголетия белков
Клетка может быстро изменять уровни белков в ответ на окружающую среду, добавляя определенные химические группы для изменения регуляции генов.
Цели обучения
Объясните, как химические модификации влияют на активность и продолжительность жизни белка
Основные выводы
Ключевые моменты
- Белки можно химически модифицировать, добавляя метильные, фосфатные, ацетильные и убиквитиновые группы.
- На продолжительность жизни белка могут влиять изменения стадий регуляции гена, включая, помимо прочего, изменение: доступность хромосомной ДНК для транскрипции, скорость трансляции, перемещение ядер, стабильность РНК и посттрансляционные модификации.
- Убиквитин добавляется к белку, чтобы пометить его для деградации протеасомой.

Ключевые термины
- убиквитин : небольшой полипептид, присутствующий в клетках всех эукариот; он участвует в модификации и разложении белков
- протеасома : сложный белок, обнаруженный в бактериальных, архейных и эукариотических клетках, который расщепляет другие белки посредством протеолиза
Химические модификации, активность белков и долговечность
Белки можно химически модифицировать путем добавления метильных, фосфатных, ацетильных и убиквитиновых групп.Добавление или удаление этих групп из белков регулирует их активность или продолжительность их существования в клетке. Иногда эти модификации могут регулировать местонахождение белка в клетке; например, в ядре, цитоплазме или прикреплены к плазматической мембране.
Иногда эти модификации могут регулировать местонахождение белка в клетке; например, в ядре, цитоплазме или прикреплены к плазматической мембране.
Химические модификации происходят в ответ на внешние раздражители, такие как стресс, недостаток питательных веществ, тепло или воздействие ультрафиолетового света. Эти изменения могут изменить функцию белка, эпигенетическую доступность, транскрипцию, стабильность мРНК или трансляцию; все это приводит к изменениям экспрессии различных генов.Это эффективный способ для клетки быстро изменить уровни содержания определенных белков в ответ на окружающую среду. Поскольку белки участвуют в каждой стадии регуляции гена, фосфорилирование белка (в зависимости от модифицированного белка) может изменить доступность хромосомы, может изменить трансляцию (путем изменения связывания или функции фактора транскрипции), может изменить перемещение ядра ( влияя на модификации комплекса ядерных пор), может изменять стабильность РНК (путем связывания или не связывания с РНК для регулирования ее стабильности), может изменять трансляцию (увеличивать или уменьшать) или может изменять посттрансляционные модификации (добавлять или удалять фосфаты или другие химические модификации). На все эти белковые активности влияет процесс фосфорилирования. Ферменты, отвечающие за фосфорилирование, известны как протеинкиназы. Добавление фосфатной группы к белку может привести либо к активации, либо к дезактивации; это зависит от белка.
На все эти белковые активности влияет процесс фосфорилирования. Ферменты, отвечающие за фосфорилирование, известны как протеинкиназы. Добавление фосфатной группы к белку может привести либо к активации, либо к дезактивации; это зависит от белка.
Другой пример химических модификаций, влияющих на активность белка, включает добавление или удаление метильных групп. Метильные группы добавляются к белкам в процессе метилирования; это наиболее распространенная форма посттрансляционной модификации.Добавление метильных групп к белку может привести к межбелковым взаимодействиям, которые позволяют регулировать транскрипцию, реагировать на стресс, репарацию белка, ядерный транспорт и даже процессы дифференцировки. Метилирование атомов азота боковой цепи считается в значительной степени необратимым, в то время как метилирование карбоксильных групп потенциально обратимо. Метилирование белков устраняет отрицательный заряд на нем и увеличивает гидрофобность белка. Метилирование боковых цепей карбоксилата покрывает отрицательный заряд и увеличивает гидрофобность.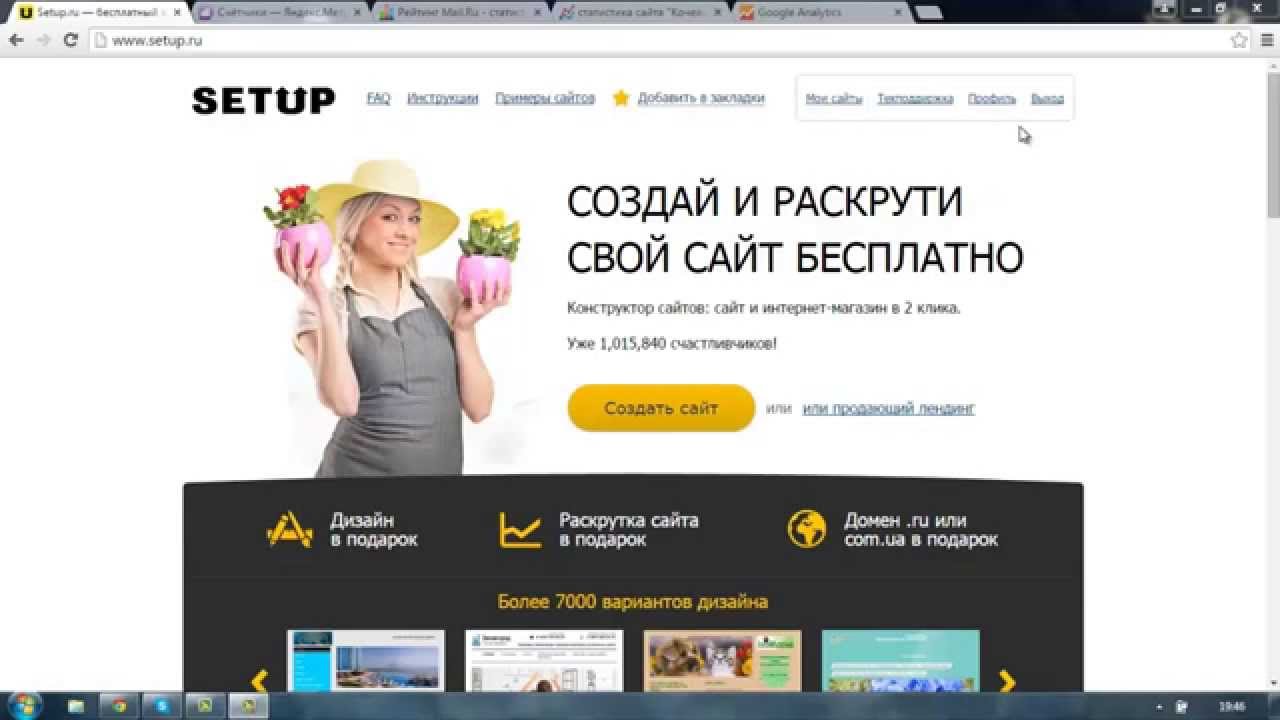 Добавление этой химической группы изменяет свойства белка и, таким образом, влияет на его активность.
Добавление этой химической группы изменяет свойства белка и, таким образом, влияет на его активность.
Добавление убиквитиновой группы к белку отмечает этот белок для разложения. Убиквитин действует как флаг, указывающий на то, что продолжительность жизни белка завершена. Эти белки перемещаются в протеасому, органеллу, которая выполняет функцию удаления белков, подлежащих разложению. Один из способов контролировать экспрессию гена — изменить продолжительность жизни белка: убиквитинирование сокращает продолжительность жизни белка.
Убиквитиновые метки : Белки с убиквитиновыми метками помечены для деградации внутри протеасомы.
Инициирование транскрипции РНК-полимеразы II: структурное представление
У эукариот РНК-полимераза II транскрибирует информационные РНК и
несколько малых ядерных РНК. Как и РНК-полимеразы I и III, полимераза
Я-я не могу действовать в одиночку. Вместо этого общие факторы инициации
[фактор транскрипции (TF) IIB, TFIID, TFIIE, TFIIF и TFIIH]
собираются на промоторной ДНК с помощью полимеразы II, создавая большой
комплекс мультипротеин-ДНК, который поддерживает точное инициирование. Другой
группа акцессорных факторов, активаторов транскрипции и
коактиваторы, регулируют скорость синтеза РНК из каждого гена в
реакция на различные сигналы развития и окружающей среды. Наш
обзор текущих знаний об этом сложном макромолекулярном механизме
подробно, с особым акцентом на идеи, полученные из структурных
исследования факторов транскрипции.
Другой
группа акцессорных факторов, активаторов транскрипции и
коактиваторы, регулируют скорость синтеза РНК из каждого гена в
реакция на различные сигналы развития и окружающей среды. Наш
обзор текущих знаний об этом сложном макромолекулярном механизме
подробно, с особым акцентом на идеи, полученные из структурных
исследования факторов транскрипции.
Эукариотическая РНК-полимераза II (pol II) представляет собой 12-субъединицу ДНК-зависимая РНК-полимераза, отвечающая за транскрипцию ядерные гены, кодирующие информационные РНК и несколько малых ядерных РНК (1).Несмотря на очевидную структурную сложность, эта мультисубъединица ферменту требуются две группы вспомогательных белков для решения двух критических биохимические проблемы. Во-первых, pol II не может распознать свою цель промоутеры напрямую. Во-вторых, pol II должен уметь модулировать производство транскриптов РНК отдельных генов в ответ на онтогенетические и сигналы окружающей среды.
Анатомия промотора и преинициативный комплекс (PIC)
Промоторы ядерных генов класса II содержат комбинации ДНК
последовательности, которые включают основные или базальные элементы промотора, промотор
проксимальные элементы и дистальные энхансерные элементы. Транскрипция
инициация pol II точно регулируется факторами транскрипции
(белки), которые взаимодействуют с этими тремя классами ДНК-мишеней и
также друг с другом (см. ссылки 2–4). Лучше всего охарактеризован
основные промоторные элементы, которые могут функционировать независимо или
синергетически это элемент ТАТА (расположенный на 25 п.н. выше
сайт начала транскрипции с консенсусной последовательностью TATAa / tAa / t), и
богатый пиримидином инициаторный элемент (расположенный в стартовом сайте). В
основной промотор представляет собой основную ДНК-мишень для pol II, и
точная инициация транскрипции зависит от сборки pol II и
факторы транскрипции (TF) IID, IIB, IIF, IIE и IIH в PIC
(Таблица 1; Рис.1). это
считали, что эти факторы транскрипции необходимы для позиционирования pol
II на большинстве промоторов ядерных генов класса II, и они обычно
называемые общими факторами инициирования (см. ссылку 2). Таким образом,
PIC функционально эквивалентен гораздо более простому Escherichia coli холоэнзим, который состоит из ядра
Субъединицы РНК-полимеразы и σ-фактор (см.
Транскрипция
инициация pol II точно регулируется факторами транскрипции
(белки), которые взаимодействуют с этими тремя классами ДНК-мишеней и
также друг с другом (см. ссылки 2–4). Лучше всего охарактеризован
основные промоторные элементы, которые могут функционировать независимо или
синергетически это элемент ТАТА (расположенный на 25 п.н. выше
сайт начала транскрипции с консенсусной последовательностью TATAa / tAa / t), и
богатый пиримидином инициаторный элемент (расположенный в стартовом сайте). В
основной промотор представляет собой основную ДНК-мишень для pol II, и
точная инициация транскрипции зависит от сборки pol II и
факторы транскрипции (TF) IID, IIB, IIF, IIE и IIH в PIC
(Таблица 1; Рис.1). это
считали, что эти факторы транскрипции необходимы для позиционирования pol
II на большинстве промоторов ядерных генов класса II, и они обычно
называемые общими факторами инициирования (см. ссылку 2). Таким образом,
PIC функционально эквивалентен гораздо более простому Escherichia coli холоэнзим, который состоит из ядра
Субъединицы РНК-полимеразы и σ-фактор (см. Ссылку 6). Промоутер
проксимальные элементы располагаются где-нибудь между 50 и 200 п.н. перед
стартовый сайт и связывание активаторов транскрипции с этими последовательностями
регулируют транскрипцию.Наконец, дистальные энхансерные элементы, которые могут быть
находится далеко от сайта инициации транскрипции в любом направлении
и ориентации, составляют еще одну группу ДНК-мишеней для факторов
модулирующий активность pol II.
Ссылку 6). Промоутер
проксимальные элементы располагаются где-нибудь между 50 и 200 п.н. перед
стартовый сайт и связывание активаторов транскрипции с этими последовательностями
регулируют транскрипцию.Наконец, дистальные энхансерные элементы, которые могут быть
находится далеко от сайта инициации транскрипции в любом направлении
и ориентации, составляют еще одну группу ДНК-мишеней для факторов
модулирующий активность pol II.
Факторы инициации транскрипции общего класса II из клетки человека
( A ) Сборка PIC начинается с TFIID
распознавание элемента ТАТА с последующим скоординированным наращиванием
TFIIB, нефосфорилированная форма pol II (pol IIA) плюс TFIIF,
TFIIE и TFIIH.Перед удлинением pol II фосфорилируется (pol IIO).
После завершения фосфатаза рециклирует pol II в свой
нефосфорилированная форма, позволяющая ферменту повторно инициировать транскрипцию in vitro . Привязка TBP (и TFIID) к блоку TATA является
по сути медленный шаг, приводящий к долгоживущему комплексу белок-ДНК. Эффективная повторная инициация транскрипции может быть достигнута, если переработанный pol
II повторно входит в преинициативный комплекс до того, как TFIID диссоциирует от
основной промоутер. (По материалам исх.5.) ( B ) Схема
представление функциональных взаимодействий, которые модулируют базальную
( Верхний ) и активатор-зависимая транскрипция
( Нижний ). Базальные факторы TBP, TFIIB, TFIIF, TFIIE и
TFIIH и pol II обозначены желтыми символами с общим
содержание фактора инициации «холофермента pol II», заключенного в
квадратных скобках. TAF II и без TAF II показаны коактиваторы (фиолетовый) и активаторы транскрипции (зеленый)
взаимодействуют со своими целями в ПОС.(Изображение предоставлено R.G.
Рёдер и С. Стивенс, Университет Рокфеллера.)
Эффективная повторная инициация транскрипции может быть достигнута, если переработанный pol
II повторно входит в преинициативный комплекс до того, как TFIID диссоциирует от
основной промоутер. (По материалам исх.5.) ( B ) Схема
представление функциональных взаимодействий, которые модулируют базальную
( Верхний ) и активатор-зависимая транскрипция
( Нижний ). Базальные факторы TBP, TFIIB, TFIIF, TFIIE и
TFIIH и pol II обозначены желтыми символами с общим
содержание фактора инициации «холофермента pol II», заключенного в
квадратных скобках. TAF II и без TAF II показаны коактиваторы (фиолетовый) и активаторы транскрипции (зеленый)
взаимодействуют со своими целями в ПОС.(Изображение предоставлено R.G.
Рёдер и С. Стивенс, Университет Рокфеллера.)
Фактор транскрипции IID
В самом общем случае производство матричной РНК начинается с
TFIID распознает и жестко связывается с элементом TATA (рис. 1).
Важная роль TFIID сделала его центром значительного
биохимические и генетические исследования с момента его открытия на клетках человека в
1980 (7). Наша текущая перепись клонированных субъединиц TFIID включает больше
более дюжины различных полипептидов массой от 15 до 250 кДа
(рассмотрено в исх.8). Большинство этих субъединиц TFIID отображают
значительная консервация среди человека, Drosophila и дрожжей,
подразумевая общий наследственный TFIID, и исследования нарушения генов четырех
субъединицы TFIID дрожжей показали, что они необходимы для жизнеспособности (9,
10).
1).
Важная роль TFIID сделала его центром значительного
биохимические и генетические исследования с момента его открытия на клетках человека в
1980 (7). Наша текущая перепись клонированных субъединиц TFIID включает больше
более дюжины различных полипептидов массой от 15 до 250 кДа
(рассмотрено в исх.8). Большинство этих субъединиц TFIID отображают
значительная консервация среди человека, Drosophila и дрожжей,
подразумевая общий наследственный TFIID, и исследования нарушения генов четырех
субъединицы TFIID дрожжей показали, что они необходимы для жизнеспособности (9,
10).
человеческим TFIID было впервые продемонстрировано с аденовирусом.
основной поздний промотор (AdMLP) (11). Исследования футпринтинга ДНКазы I
AdMLP и промоторы отдельных генов человека выявили специфичные для последовательности
взаимодействия между человеческим TFIID и элементом TATA, которые
в первую очередь опосредуется субъединицей TBP TFIID (см. ниже).В
Напротив, защита как перед, так и после элемента TATA
в значительной степени независима от последовательности, демонстрирует нуклеосомоподобный образец
Гиперчувствительность к ДНКазе I, радикально варьирует между промоторами и может
индуцироваться некоторыми активаторами (см. ссылку 8). Это замечательно
что связывание коробки TATA с помощью TFIID или TBP препятствует упаковке
коровой промотор с несвязными гистоновыми белками (h3A, h3B, h4 и
h5). Напротив, упаковка ядерного промотора октамерами гистонов в
нуклеосомы предотвращают связывание TFIID или TBP с элементом TATA,
эффективно подавляя транскрипцию (см.12). В
vivo , репрессия транскрипции, опосредованная хроматином, преодолевается
различные АТФ-зависимые макромолекулярные машины (например, SWI / SNF
комплекс), которые ремоделируют хроматин в непосредственной близости от корового промотора
(рассмотрено в ссылке 13).
ссылку 8). Это замечательно
что связывание коробки TATA с помощью TFIID или TBP препятствует упаковке
коровой промотор с несвязными гистоновыми белками (h3A, h3B, h4 и
h5). Напротив, упаковка ядерного промотора октамерами гистонов в
нуклеосомы предотвращают связывание TFIID или TBP с элементом TATA,
эффективно подавляя транскрипцию (см.12). В
vivo , репрессия транскрипции, опосредованная хроматином, преодолевается
различные АТФ-зависимые макромолекулярные машины (например, SWI / SNF
комплекс), которые ремоделируют хроматин в непосредственной близости от корового промотора
(рассмотрено в ссылке 13).
ТАТА-бокс-связывающий белок
После публикации последовательности дрожжевого ТВР в 1989 г.
быстро последовательностями гомологичных генов от различных эукариот
и архебактерии (идентичность аминокислот в пределах
филогенетически консервативная часть из 180 остатков составляет от 38% до
100%, рассмотрено в исх.14). Один рекомбинантный ТВР может связывать оба
общие и нормативные факторы и прямая сборка ПОС в
vitro и базальная транскрипция (см. ссылку 2). Базальный или стержневой
промотор-зависимая транскрипция является относительно неэффективной в
vitro , которая послужила важным инструментом для
характеризуя минимальные требования Pol II к TBP, TFIIB, TFIIF,
TFIIE, и TFIIH и центральная область промотора сразу
перед сайтом начала транскрипции. Помимо определения
минимальные факторы, необходимые для точного инициирования Pol II, базальная
система транскрипции была использована для установления порядка PIC
сборка, изображенная на рис.1 А .
Рис. 1 B противопоставляет базальную и активированную транскрипцию
(рассмотрено в ссылке 8). Активированная транскрипция in vivo требуется весь промотор, который включает центральную область плюс
проксимальная и дистальная области энхансера промотора. Транскрипционный
активаторы регулируют эффективность инициации pol II, распознавая
их последовательности ДНК-мишени в проксимальном или дистальном энхансере промотора
элементы и взаимодействие с PIC (возможно, через посредников
известные как коактиваторы). Для активированной транскрипции требуется TBP и оставшиеся субъединицы TFIID (
TAF II s), другие общие факторы инициации TFIIB, TFIIF,
TFIIE и TFIIH, а также активаторы и коактиваторы транскрипции.
Для активированной транскрипции требуется TBP и оставшиеся субъединицы TFIID (
TAF II s), другие общие факторы инициации TFIIB, TFIIF,
TFIIE и TFIIH, а также активаторы и коактиваторы транскрипции.
Трехмерная структура консервативной части ТВР
поразительно похожий на седло (14–16) (рис.
2 A ), что идеально соответствует
Биохимическая функция ТВР как белка, который сидит на ДНК, создавая
стабильная платформа для связывания других факторов транскрипции.Связывание ДНК
поддерживается вогнутой нижней стороной седла, в то время как выпуклая
верхняя поверхность или посадочное место седла связывает различные компоненты
аппарат транскрипции (см. ссылку 14). ТБП состоит из двух
квазиидентичные домены (рис. 2 A ), соответствующие
два прямых повтора, обнаруженные в консервативной части ТВР (15).
Следовательно, предок ТВР мог функционировать как димер с геном
дупликация и слияние, приводящее к мономерному квазисимметричному ТВР.
( A ) Объемные конструкции
TBP (14–16) ( верхний левый ), TBP в комплексе с TATA
элемент (17–19) ( Правый верхний ), клемма C или сердцевина TFIIB
(cTFIIB) –TBP – TATA элементный тройной комплекс (20) ( Нижний
Слева ) и тройной элемент TFIIA – TBP – TATA (21, 22)
( Нижний правый ).Белки изображены в виде ленты.
рисунки, с маркированными концами N и C, если они видны. ДНК — это
показаны в виде фигурки с гипотетическими линейными удлинениями B-формы
на обоих концах. Сайт начала транскрипции AdMLP помечен
+1. TBP, а комплексы TBP – ДНК и cTFIIB – TBP – ДНК показаны из
та же самая выгодная точка после сайта начала транскрипции. В
Комплекс TFIIA – TBP – ДНК рассматривается выше элемента TATA,
смотрит в сторону начала транскрипции.Молекулы имеют цветовую маркировку
следующим образом: красный, первый повтор cTFIIB; пурпурный, второй повтор cTFIIB;
светло-голубой, конец TBP N и первый повтор; темно-синий, секунда TBP
повторить; зеленый — малая субъединица TFIIA; желтый — большая субъединица TFIIA; а также
серый, ДНК. Когда TBP распознает малую бороздку элемента TATA,
ДНК изгибается и разматывается, чтобы представить края малых бороздок
основания к нижней стороне молекулярного седла. На cTFIIB или TFIIA
связывания с комплексом ТВР-ДНК практически нет изменений в
структура бинарного комплекса.( B ) Детали конструкции
TFIIB. Относительная ориентация двух доменов cTFIIB в
свободная и связанная форма совершенно разные. Связанные и свободные cTFIIB
нарисованы с выровненными первыми доменами. Концы N и C
фрагменты белка, использованные в структурных исследованиях, помечены, а
α-спирали каждого домена cTFIIB окрашены в красный, зеленый,
синий, желтый и пурпурный. Спираль присутствует только во второй области
cTFIIB в тройном комплексе окрашен в голубой цвет.Структура
cTFIIB в тройном комплексе cTFIIB – TBP – TATA элемент (20)
( Левый ). Структура свободного cTFIIB (23)
( Центр ). Структура N-конца, Zn 2+ связывающая область TFIIB (24) ( Right ).
Когда TBP распознает малую бороздку элемента TATA,
ДНК изгибается и разматывается, чтобы представить края малых бороздок
основания к нижней стороне молекулярного седла. На cTFIIB или TFIIA
связывания с комплексом ТВР-ДНК практически нет изменений в
структура бинарного комплекса.( B ) Детали конструкции
TFIIB. Относительная ориентация двух доменов cTFIIB в
свободная и связанная форма совершенно разные. Связанные и свободные cTFIIB
нарисованы с выровненными первыми доменами. Концы N и C
фрагменты белка, использованные в структурных исследованиях, помечены, а
α-спирали каждого домена cTFIIB окрашены в красный, зеленый,
синий, желтый и пурпурный. Спираль присутствует только во второй области
cTFIIB в тройном комплексе окрашен в голубой цвет.Структура
cTFIIB в тройном комплексе cTFIIB – TBP – TATA элемент (20)
( Левый ). Структура свободного cTFIIB (23)
( Центр ). Структура N-конца, Zn 2+ связывающая область TFIIB (24) ( Right ).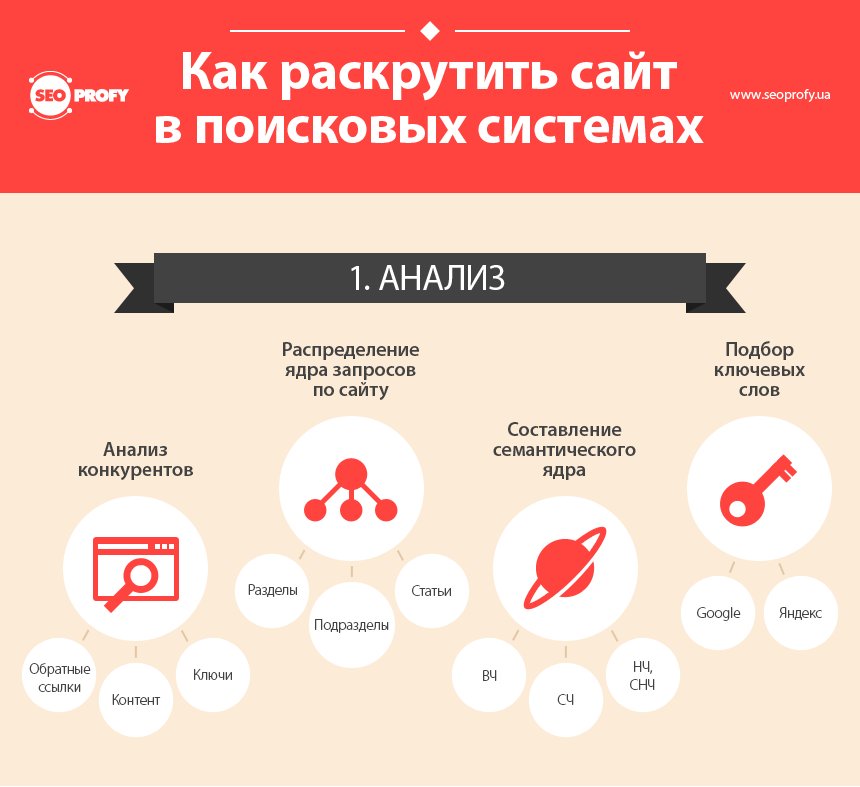 Атом Zn
окрашен в красный цвет. 60 остатков между С-концом
Связывающий домен Zn 2+ и N-конец cTFIIB являются
гибкие и не визуализировались в структурных
исследования.
Атом Zn
окрашен в красный цвет. 60 остатков между С-концом
Связывающий домен Zn 2+ и N-конец cTFIIB являются
гибкие и не визуализировались в структурных
исследования.
Структуры ТБФ растений (17, 25), дрожжей (18) и человека (19) в комплексе
с различными элементами ТАТА (рис.2 А ). Эти три сокристаллические структуры очень
похожи и демонстрируют общий механизм индуцированной подгонки (26)
белок-ДНК узнавание (G. Patikoglou, J. L. Kim, S.K.B.,
неопубликованные данные). Связывание ДНК опосредуется изогнутыми антипараллельными
β-лист, который обеспечивает большую вогнутую поверхность для малых канавок и
Магистральные контакты с элементом ТАТА размером 8 п.н. 5 ‘конец стандарта
B-форма ДНК входит в нижнюю часть молекулярного седла, где TBP
производит резкий переход к беспрецедентному, частично раскрученному
форма правой двойной спирали, индуцированной вставкой двух
остатки фенилаланина в первую стадию Т: A основания.После этого
расширенная поверхность малой бороздки размотанной, плавно изогнутой ДНК
приближается к нижней стороне молекулярного седла, что позволяет
прямые взаимодействия между боковыми цепями белка и малой бороздкой
края центральной 6 п. н. Второй большой перегиб вызван вставкой
двух остатков фенилаланина в основную стадию между двумя последними
пары оснований элемента TATA, и имеется соответствующий резкий
вернуться к B-форме ДНК. Несмотря на это огромное искажение, Уотсон – Крик
спаривание оснований сохраняется повсюду, и, похоже, нет
спиральная деформация, индуцированная в ДНК из-за частичного раскручивания.
компенсируется правосторонней сверхспирализацией двойной спирали.
н. Второй большой перегиб вызван вставкой
двух остатков фенилаланина в основную стадию между двумя последними
пары оснований элемента TATA, и имеется соответствующий резкий
вернуться к B-форме ДНК. Несмотря на это огромное искажение, Уотсон – Крик
спаривание оснований сохраняется повсюду, и, похоже, нет
спиральная деформация, индуцированная в ДНК из-за частичного раскручивания.
компенсируется правосторонней сверхспирализацией двойной спирали.
Упаковка ДНК в нуклеосомы включает в себя наматывание двойной спирали вокруг
октамер гистонов. Последовательно-зависимое позиционирование нуклеосом
коррелирует с изгибом A + T-богатых последовательностей к малой бороздке (28,
29), и упаковка элементов ТАТА в нуклеосомы, вероятно, приводит к
при сжатии малой канавки, исключающей связывание ТВР. Наоборот, a
предварительно сформированный PIC остается транскрипционно активным после нуклеосомы
сборки (30), и только рекомбинантный дрожжевой TBP предотвращает
репрессия транскрипции, опосредованная нуклеосомами (31). Таким образом
сокристаллические структуры комплексов ТВР-ДНК могут обеспечить простой
механическое объяснение взаимного исключения упаковки ДНК и
транскрипция. Широко распространено мнение, что активаторы транскрипции
связаны с промотором проксимальных и / или дистальных энхансерных элементов-мишеней
факторы ремоделирования нуклеосом для основных промоторов генов, предназначенных для
выражение. После ремоделирования хроматина TFIID сможет
распознать элемент TATA и начать сборку PIC (см.
32).
Таким образом
сокристаллические структуры комплексов ТВР-ДНК могут обеспечить простой
механическое объяснение взаимного исключения упаковки ДНК и
транскрипция. Широко распространено мнение, что активаторы транскрипции
связаны с промотором проксимальных и / или дистальных энхансерных элементов-мишеней
факторы ремоделирования нуклеосом для основных промоторов генов, предназначенных для
выражение. После ремоделирования хроматина TFIID сможет
распознать элемент TATA и начать сборку PIC (см.
32).
под действием TBP также может иметь значение для координации и / или стабилизация сборки ПОС и взаимодействия активатор-ПОС. Сборка PIC вокруг изгиба может образовывать более компактный комплекс мультипротеин-ДНК. Более того, изгиб ДНК под действием TBP может помочь в образовании петли ДНК, чтобы привести удаленно связанные активаторы транскрипции ближе к коровому промотору для взаимодействия с компонентами ПОС.
Дополнительные биофизические методы были использованы для изучения взаимодействий
между TBP и ДНК. Эксперименты по выбору сайта с Acanthamoeba TBP продемонстрировала явное предпочтение
аналогично изученным кристаллографически (33). Изгиб ДНК с помощью TBP
в растворе было подтверждено с помощью анализа круговой перестановки (34). TBP
также было показано, что связывание усиливается за счет предварительного изгиба ДНК по направлению к
большая канавка и подавляется предварительным изгибом к малой канавке (35).
Кинетика ассоциации ТВР – ДНК изучалась различными методами.
(36–38), которые дали результаты, согласующиеся с формированием начального
комплекс столкновения с последующей медленной стадией изомеризации с
константа скорости второго порядка около 10 6 M -1 · с -1 .После формирования окно TBP – TATA
комплекс очень стабилен и измеренный период полужизни дрожжей
Время действия комплекса ТБФ – AdMLP в водном растворе составляет примерно 2 ч (36).
Наконец, новое исследование химической модификации продемонстрировало, что ядро
искажение промотора временно выходит за пределы 3′-конца TATA
элемент во время связывания ТВР (39).
Эксперименты по выбору сайта с Acanthamoeba TBP продемонстрировала явное предпочтение
аналогично изученным кристаллографически (33). Изгиб ДНК с помощью TBP
в растворе было подтверждено с помощью анализа круговой перестановки (34). TBP
также было показано, что связывание усиливается за счет предварительного изгиба ДНК по направлению к
большая канавка и подавляется предварительным изгибом к малой канавке (35).
Кинетика ассоциации ТВР – ДНК изучалась различными методами.
(36–38), которые дали результаты, согласующиеся с формированием начального
комплекс столкновения с последующей медленной стадией изомеризации с
константа скорости второго порядка около 10 6 M -1 · с -1 .После формирования окно TBP – TATA
комплекс очень стабилен и измеренный период полужизни дрожжей
Время действия комплекса ТБФ – AdMLP в водном растворе составляет примерно 2 ч (36).
Наконец, новое исследование химической модификации продемонстрировало, что ядро
искажение промотора временно выходит за пределы 3′-конца TATA
элемент во время связывания ТВР (39).
Фактор транскрипции IIB
TFIIB — следующий общий фактор инициации для входа в PIC. В
образовавшаяся платформа TFIIB-TFIID-ДНК, в свою очередь, распознается комплексом
pol II и TFIIF, за которыми следуют TFIIE и TFIIH (рис.1). В
vitro исследования с отрицательно свернутым геном иммуноглобулина
промотор продемонстрировал, что точное инициирование транскрипции может быть
восстановлен с помощью TBP, TFIIB и pol II, что позволяет предположить, что вместе TBP
и положение pol II TFIIB (40). Предположительно, энергия, обеспечиваемая
отрицательная суперспирализация способствует плавлению промотора на
сайт начала транскрипции, которому обычно способствует
АТФ-зависимая субъединица ДНК-геликазы TFIIH (см. Ниже). Мутации в
TFIIB изменяет стартовые сайты pol II в дрожжах, как и мутации в большом
субъединицы pol II, что дает убедительные доказательства ее функции в качестве
точный спейсер / мостик между TFIID и pol II на промоторе ядра
который определяет сайт начала транскрипции (см. 20).
20).
Второй этап сборки PIC также оказался поддающимся рентгеновскому облучению. кристаллографическое исследование. Структура элемента TFIIB – TBP – TATA. тройной комплекс был описан в 1995 г. (20) (рис. 2 A ). С-концевой или сердцевинный TFIIB (cTFIIB) представляет собой двухдоменный α-спиральный белок. который является структурным гомологом белка клеточного цикла циклина А (41, 42) (рис.2 B ). Несмотря на эту замечательную конструктивную сходства, нет доказательств того, что TFIIB регулирует активность любая циклин-зависимая киназа.Более того, наличие пара циклин / циклин-зависимая киназа в TFIIH, по-видимому, вероятность того, что TFIIB будет иметь циклиноподобное поведение, маловероятна.
Тройной комплекс cTFIIB – TBP – ДНК образуется cTFIIB
зажимание кислого С-концевого стремени ТВР в его основной щели, и
взаимодействуя с основной цепью фосфорибозы до и после
центр элемента ТАТА. Первый домен cTFIIB образует
нижележащая поверхность тройного комплекса cTFIIB – TBP – ДНК, где
вместе с N-концевым доменом TFIIB (24) (проиллюстрировано на рис. 2 B ) он легко может действовать как мост между TBP и
pol II, чтобы исправить сайт начала транскрипции. Остальные
доступные для растворителя поверхности TBP и TFIIB обширны,
обеспечение достаточного количества сайтов узнавания для связывания TAF II s,
другие факторы инициации класса II, активаторы транскрипции и
соактиваторы. Сама структура комплекса элементов ТВП – ТАТА имеет следующий вид:
по существу не изменяется тройным комплексообразованием. cTFIIB
распознает предварительно собранный комплекс ТВР-ДНК, включая путь
каркас фосфорибозы, созданный беспрецедентной деформацией ДНК
индуцируется связыванием ТВР.В дополнение к стабилизации TBP – ДНК
комплекса, связывание TFIIB может вносить вклад в полярность элемента ТАТА
признание. Если бы TBP связывался с квазисимметричным блоком TATA в
неправильная ориентация (т.е. если N-концевая половина молекулярного седла
должны были взаимодействовать с 5 ‘концом элемента TATA),
основная / гидрофобная поверхность N-концевого хомута сделает
неблагоприятные электростатические взаимодействия с основной щелью TFIIB.
2 B ) он легко может действовать как мост между TBP и
pol II, чтобы исправить сайт начала транскрипции. Остальные
доступные для растворителя поверхности TBP и TFIIB обширны,
обеспечение достаточного количества сайтов узнавания для связывания TAF II s,
другие факторы инициации класса II, активаторы транскрипции и
соактиваторы. Сама структура комплекса элементов ТВП – ТАТА имеет следующий вид:
по существу не изменяется тройным комплексообразованием. cTFIIB
распознает предварительно собранный комплекс ТВР-ДНК, включая путь
каркас фосфорибозы, созданный беспрецедентной деформацией ДНК
индуцируется связыванием ТВР.В дополнение к стабилизации TBP – ДНК
комплекса, связывание TFIIB может вносить вклад в полярность элемента ТАТА
признание. Если бы TBP связывался с квазисимметричным блоком TATA в
неправильная ориентация (т.е. если N-концевая половина молекулярного седла
должны были взаимодействовать с 5 ‘концом элемента TATA),
основная / гидрофобная поверхность N-концевого хомута сделает
неблагоприятные электростатические взаимодействия с основной щелью TFIIB.
Также была определена структура ЯМР раствора одного cTFIIB. (Рисунок.2 В ) (23). Хотя каждый домен в ЯМР структура очень похожа на своего аналога в рентгеновской структуре, две структуры демонстрируют разное пространственное расположение два домена. Эти данные предполагают, что олигопептидный линкер между два домена являются гибкими, и этот TFIIB претерпевает конформационную изменение при распознавании преформированного комплекса ТВР – ДНК. Таким образом, TFIIB, как и TBP распознает свою цель через индуцированную подгонку (Г. Патикоглу, Дж. Л. Ким, и С.К.Б., неопубликованные данные).
Факторы транскрипции IIE, -IIF и -IIH
После образования комплекса TFIIB – TFIID – ДНК три других
общие факторы инициации и pol II завершают растущий PIC. ТФИИФ
представляет собой гетеродимер субъединиц массой 30 и 74 кДа (см. обзор в
исх. 43). Среди общих инициирующих факторов TFIIF уникален своим
способность образовывать очень стабильный комплекс с pol II, называемый
pol / F (рис. 1). Хотя нет структурного
информация, доступная для TFIIF, результаты сайт-ориентированных
мутагенез и исследования сшивания белок-ДНК дают некоторые
информация о его местонахождении в пределах ПОС.Аланин-сканирование
мутагенез ТВР человека выявил единственный остаток, необходимый для TFIIF
связка, которая расположена на выпуклой верхней поверхности молекулярного
седло на его выходной стороне (44). Фотосшивающие исследования
идентифицировали перекрестные связи между субъединицами TFIIF и положениями -5,
−15 и −19 (45). Вместе эти данные локализуют TFIIF внутри PIC.
к области основного промотора между 3′-концом ТАТА-бокса
(позиция -24) и сайт старта транскрипции (рис.
2 А ).TFIIE представляет собой α 2 β 2 гетеротетрамер субъединиц массой 34 и 56 кДа (обзор в
исх. 46). Исследования фотоперекрестного сшивания выявили сшивки между
Субъединица TFIIE массой 34 кДа и положения -2 и -14 (45), которые локализуют
TFIIE к той же части основного промотора, что и TFIIF.
1). Хотя нет структурного
информация, доступная для TFIIF, результаты сайт-ориентированных
мутагенез и исследования сшивания белок-ДНК дают некоторые
информация о его местонахождении в пределах ПОС.Аланин-сканирование
мутагенез ТВР человека выявил единственный остаток, необходимый для TFIIF
связка, которая расположена на выпуклой верхней поверхности молекулярного
седло на его выходной стороне (44). Фотосшивающие исследования
идентифицировали перекрестные связи между субъединицами TFIIF и положениями -5,
−15 и −19 (45). Вместе эти данные локализуют TFIIF внутри PIC.
к области основного промотора между 3′-концом ТАТА-бокса
(позиция -24) и сайт старта транскрипции (рис.
2 А ).TFIIE представляет собой α 2 β 2 гетеротетрамер субъединиц массой 34 и 56 кДа (обзор в
исх. 46). Исследования фотоперекрестного сшивания выявили сшивки между
Субъединица TFIIE массой 34 кДа и положения -2 и -14 (45), которые локализуют
TFIIE к той же части основного промотора, что и TFIIF. TFIIH — это
большая мультипротеиновая сборка, состоящая из девяти субъединиц, которые варьируются в
масса от 39 до 89 кДа (см. ссылку 47). В отличие от другого генерала
факторы инициации, TFIIH поддерживает различную каталитическую активность,
включая ДНК-зависимую АТФазу, АТФ-зависимую ДНК-геликазу и
серин / треонинкиназа, способная фосфорилировать
С-концевой домен большой субъединицы pol II и регулируется
субъединица H циклина.По крайней мере, две субъединицы TFIIH (ERCC2 и
ERCC3) также являются компонентами механизма эксцизионной репарации ДНК, который
предполагает, что мультипротеиновый комплекс TFIIH может также участвовать в
Ремонт ДНК (см. Ссылку 48).
TFIIH — это
большая мультипротеиновая сборка, состоящая из девяти субъединиц, которые варьируются в
масса от 39 до 89 кДа (см. ссылку 47). В отличие от другого генерала
факторы инициации, TFIIH поддерживает различную каталитическую активность,
включая ДНК-зависимую АТФазу, АТФ-зависимую ДНК-геликазу и
серин / треонинкиназа, способная фосфорилировать
С-концевой домен большой субъединицы pol II и регулируется
субъединица H циклина.По крайней мере, две субъединицы TFIIH (ERCC2 и
ERCC3) также являются компонентами механизма эксцизионной репарации ДНК, который
предполагает, что мультипротеиновый комплекс TFIIH может также участвовать в
Ремонт ДНК (см. Ссылку 48).
Шаги сборки PIC, описанные выше, были установлены в
vitro с использованием минимальной системы транскрипции, изображенной на рис.
1 B Верхний . Они не обязательно являются единственным средством
из которых можно собрать функциональную ПОС. В последнее время ряд крупных
мультипротеиновые комплексы, содержащие pol II и большую часть общих
факторы инициации (кроме TFIID и TFIIB) плюс комплекс SRB
и другие белки были очищены из ядерного экстракта (см. исх.49). Такие комплексы обычно называют «pol II.
голоферменты », что не совсем правильно, потому что они не могут
функционировать в одиночку. Эти захватывающие открытия предполагают, что в
vivo PIC можно собрать всего за несколько шагов (например, TFIID
плюс ДНК с последующим добавлением TFIIB и затем «pol II
холоэнзим », как показано на фиг. 1 B ).
исх.49). Такие комплексы обычно называют «pol II.
голоферменты », что не совсем правильно, потому что они не могут
функционировать в одиночку. Эти захватывающие открытия предполагают, что в
vivo PIC можно собрать всего за несколько шагов (например, TFIID
плюс ДНК с последующим добавлением TFIIB и затем «pol II
холоэнзим », как показано на фиг. 1 B ).
Цикл инициации транскрипции РНК Pol II
После завершения сборки PIC и в присутствии нуклеозида
трифосфаты, происходит разделение цепи в месте старта транскрипции
чтобы дать открытый комплекс, C-концевой домен большой субъединицы
pol II фосфорилируется (предположительно киназной субъединицей TFIIH),
а pol II инициирует транскрипцию и высвобождается с промотора.Во время удлинения in vitro TFIID может оставаться связанным с
основной промотор, поддерживающий повторную инициацию транскрипции с помощью pol II и
другие общие факторы инициирования (рис. 1 A ;
рассмотрено в исх. 5). Поскольку связывание основного промотора субъединицей ТВР
TFIID — медленный шаг, цикл транскрипции
проиллюстрировано на рис. 1 A может представлять механизм
инициации pol II in vivo . И потребность в хроматине
ремоделирование, которое требует АТФ, и медленная стадия изомеризации во время
Деформация ДНК, индуцированная ТВР, будет амортизироваться при многократном инициировании
события, если TFIID оставался стабильно связанным с основным промотором
между последовательными раундами транскрипции.Этот сценарий
особенно привлекателен в контексте сокращенной сборки PIC
механизм с участием «холофермента pol II».
5). Поскольку связывание основного промотора субъединицей ТВР
TFIID — медленный шаг, цикл транскрипции
проиллюстрировано на рис. 1 A может представлять механизм
инициации pol II in vivo . И потребность в хроматине
ремоделирование, которое требует АТФ, и медленная стадия изомеризации во время
Деформация ДНК, индуцированная ТВР, будет амортизироваться при многократном инициировании
события, если TFIID оставался стабильно связанным с основным промотором
между последовательными раундами транскрипции.Этот сценарий
особенно привлекателен в контексте сокращенной сборки PIC
механизм с участием «холофермента pol II».
Регуляция инициации транскрипции РНК Pol II
Регуляция транскрипции ядерного гена класса II в
реакция на сигналы развития или окружающей среды достигается за счет
контроль сборки PIC или каталитической эффективности pol II
во время инициации, удлинения или прекращения. Когда транскрипционный
активаторы взаимодействуют с TAF II с, усиленный набор
и / или наблюдается стабилизация TFIID на промоторе (см. в исх.8). Результаты исследований с гибридными белками, состоящими из
TBP, слитый с гетерологичными ДНК-связывающими доменами, предполагает, что TFIID
привлечение к промотору может быть этапом, ограничивающим скорость (50–52), который
преодолевается взаимодействиями активатор – TAF II . В
vivo отпечаток проксимальных областей промотора некоторых
специфические для печени гены продемонстрировали, что многие транскрипционные
активаторы, по-видимому, связываются одновременно (53), что согласуется с
с точки зрения того, что два или более активатора могут оказывать синергетический эффект
по транскрипции через согласованное взаимодействие с несколькими
компоненты ПОС.Тянь и его коллеги (54, 55) недавно
предоставили прямую поддержку этой гипотезе, продемонстрировав, что
синергия между двумя разными активаторами (Bicoid и Hunchback) связана
к тем же результатам промотора, по крайней мере частично, из определенных
взаимодействия с двумя разными Drosophila TAF II s
которые увеличивают набор сотрудников TFIID.
в исх.8). Результаты исследований с гибридными белками, состоящими из
TBP, слитый с гетерологичными ДНК-связывающими доменами, предполагает, что TFIID
привлечение к промотору может быть этапом, ограничивающим скорость (50–52), который
преодолевается взаимодействиями активатор – TAF II . В
vivo отпечаток проксимальных областей промотора некоторых
специфические для печени гены продемонстрировали, что многие транскрипционные
активаторы, по-видимому, связываются одновременно (53), что согласуется с
с точки зрения того, что два или более активатора могут оказывать синергетический эффект
по транскрипции через согласованное взаимодействие с несколькими
компоненты ПОС.Тянь и его коллеги (54, 55) недавно
предоставили прямую поддержку этой гипотезе, продемонстрировав, что
синергия между двумя разными активаторами (Bicoid и Hunchback) связана
к тем же результатам промотора, по крайней мере частично, из определенных
взаимодействия с двумя разными Drosophila TAF II s
которые увеличивают набор сотрудников TFIID.
В своей простейшей форме белок-белковые взаимодействия, регулирующие pol II активность вовлекает компоненты преинициативного комплекса (TBP, TAF II s, TFIIB, pol II, TFIIF, TFIIE и TFIIH) и активаторы транскрипции (связанные либо с проксимальным, либо с дистальным промотором). энхансерные элементы).Наша текущая картина активатора – TFIID взаимодействий предполагает, что TAF II s можно рассматривать как большой мультипротеиновый комплекс, который находится поверх TBP и интегрирует сигналы из многих активаторов и соактиваторов, не относящихся к TAF II . В оставшиеся общие факторы инициации и pol II представляют разные мишени в PIC для взаимодействия с транскрипционными активаторы. Действительно, кажется вероятным, что каждый компонент ПОС мишень хотя бы одного активатора транскрипции во время транскрипция одного или нескольких из примерно 100000 классов II промоторы ядерных генов.Косвенное взаимодействие между PIC и активаторы транскрипции, опосредованные не-TAF II соактиваторы также наблюдались (рис. 1 B ). Коактиваторы, такие как человеческий PC4, человеческий OCA-B и дрожжевой SRB комплекс, может служить переходником между активаторами и базальными факторами (рассмотрено в ссылке 8).
Фактор транскрипции IIA
TFIIA был впервые описан как общий фактор инициации (7) и изначально считалось необходимым для транскрипции многих, если не все промоторы ядерных генов класса II.После обширных механистическая характеристика и клонирование генов, кодирующих субъединицы TFIIA, однако теперь ясно, что TFIIA лучше всего определяется как коактиватор, поддерживающий регуляцию транскрипции pol II (рассмотрено в ссылке 3). На ранних этапах сборки PIC TFIIA может связываться с и стабилизировать комплексы TFIID – ДНК или TFIIB – TFIID – ДНК, позволяя им отразить пагубные эффекты подавляющего отрицательные кофакторы и усиливают стимулирующее действие активаторы транскрипции (см.56).
Недавно структура тройного комплекса TFIIA – TBP – TATA element определено методом рентгеновской кристаллографии (21, 22) (рис. 2 А ). TFIIA дрожжей состоит из двух α / β субъединицы 14 и 32 кДа, которые образуют интимный гетеродимер через 12-ниточная β-цилиндрическая структура. Тройной комплекс образован TFIIA распознает N-концевую скобу TBP и взаимодействует с остов фосфорибозы перед элементом ТАТА на противоположной стороне грань двойной спирали cTFIIB (рис.2 А ). В виде в комплексе cTFIIB – TBP – ДНК TFIIA распознает предварительно сформированные Комплекс TBP – ДНК, объясняющий стабилизацию комплекса TFIID – ДНК с помощью TFIIA.
Когда структуры cTFIIB – TBP – ДНК и TFIIA – TBP – ДНК комплексы объединяются для создания модели TFIIA – TFIIB – TBP – ДНК четвертичный комплекс (рис. 3), механизм по которые TFIIB и TFIIA действуют синергетически в стабилизации TFIID-ДНК комплекс можно рационализировать. Вместо того, чтобы взаимодействовать друг с другом непосредственно основные поверхности TFIIB и TFIIA контактируют с отрицательно заряженный остов фосфорибозы на противоположных сторонах двойная спираль непосредственно перед элементом ТАТА.Модель Комплекс TFIIA – TFIIB – TBP – ДНК также дает критическую информацию о роль TFIIA как коактиватора или моста между транскрипционными активаторы и ПОС. Обе субъединицы TFIIA образуют вышестоящий поверхность тройного комплекса TFIIA – TBP – ДНК, где они находятся доступны для взаимодействия с активаторами транскрипции, связанными с промоторные проксимальные или дистальные энхансерные элементы. Следовательно, это не удивительно, что остатки на поверхности ТВР, которые участвуют в контакты с TFIIA необходимы для активации транскрипции в vivo (56, 57).
Модель комплекса TFIIA – TFIIB – TBP – ДНК на основе на структуру элемента cTFIIB – TBP – TATA (20), а Комплексы TFIIA – TBP – TATA элемент (21, 22) (см. Рис. 2 А ). Сайт начала транскрипции помечен +1. Схема цветового кодирования такая же, как на рис. 2 A . ( Верхний ) Если смотреть по оси TBP приблизительного внутримолекулярная симметрия сверху седла. ( Нижний ) Вид снизу на молекулярное седло.
Выводы и перспективы
Прошло более четверти века с тех пор, как сложность транскрипции эукариот впервые было обнаружено благодаря открытию Редера трех РНК-полимераз (58).С тех пор технически сложно биохимическая работа и элегантные генетические исследования определили и функционально охарактеризованы многие компоненты, которые вместе облегчают и регулируют производство Pol II матричной РНК. Трехмерные структуры ТВП и его комплекса с ядром промотор, cTFIIB, TFIIBn и TFIIB – TBP – ДНК и TFIIA – TBP – ДНК тройные комплексы выявили новые взаимодействия белок-ДНК, и детальная механистическая оценка того, как эти полипептиды поддерживают инициация транскрипции.Структурные биологи сейчас занимаются даже более крупные сборки факторов транскрипции, и есть все основания для полагаем, что скоро мы увидим структуры TFIIE, TFIIF, TFIID, TFIIH, и РНК pol II. Биологи факторов транскрипции в настоящее время руководят их усилия по решению проблемы понимания того, как транскрипция инициация контролируется на уровне отдельного гена. Там есть значительные доказательства того, что PIC и активаторы транскрипции и коактиваторы могут собираться на промоторе в стереоспецифический нуклеопротеидный комплекс или «транскриптосома», поддерживающий активация транскрипции (см.59).
Другой важной проблемой, которую необходимо решить, является необходимость понять сложное взаимодействие между упаковкой ДНК и транскрипция. Неожиданно недавние кристаллографические исследования задокументированные прямые структурные связи между факторами транскрипции и гистоновые белки (рис. 4), предполагая, что макромолекулярные машины, отвечающие за упаковку ДНК и транскрипция на каком-то уровне эволюционно связана. В структурные взаимосвязи, показанные на рис.4 также вызывают интригующие вопросы, касающиеся механизмов, с помощью которых гистоноподобная транскрипция факторы работают (см. ссылку 64). TFIID может содержать TAF II субструктура, которая напоминает октамер гистонов и опосредует некоторые неспецифические взаимодействия TFIID с ДНК (11). Прямое свидетельство обертывания ДНК вокруг TFIID было получено Roeder и соавторы (65), которые продемонстрировали TAF II -ДНК сшивает непосредственно перед элементом ТАТА и ниже по потоку элемент ТАТА, простирающийся в 5′-нетранслируемую область гена, и TFIID-индуцированная суперспирализация ДНК замкнутой кольцевой плазмиды.В напротив, связывание ТВР с той же плазмидой не изменяет связывание число, потому что суперспирализация ДНК TBP компенсируется частичной раскручивание двойной спирали (см. ссылку 25). Наконец, структурное сходство ядерного фактора гепатоцитов (HNF) -3γ и гистон H5 может иметь функциональное значение. Связывание HNF-3 с двумя соседние высокоаффинные сайты в энхансере гена сывороточного альбумина мыши (66), как было показано, индуцируют фазирование расположения нуклеосом. внутри энхансера (27).
Структурное сходство между транскрипцией факторы и гистоновые белки. ( верхний ) гетеротетрамерный сборка N-концевых частей двух Drosophila TAF II с (dTAF II 42 / dTAF II 62) 2 (60) и соответствующий вид гетеротетрамера гистона h4 / h5, производного из структуры октамера гистонов (61) (дополнительная N-концевая спираль h4, визуализированная в этом исследовании, была опущена для ясность).( Нижний ) ДНК-связывающий домен гепатоцита ядерный фактор-3γ (62), и соответствующий вид глобулярной домен линкерного гистона H5 (GH5) (63).
Благодарности
Мы благодарим докторов наук. Г. Аренц, С. Бэгби, Дж. Гейгер, М. Икура, Дж. Л. Ким, Э. Н. Мудрианакису, П. Б. Сиглеру, М. Саммерсу и X. Се за помощь с фигурная подготовка. Эта работа была поддержана Ховардом Хьюзом. Медицинский институт (S.K.B.) и выпускник Университета Рокфеллера Товарищество (Д.Б.Н.).
Сноски
↵ Текущий адрес: Клеточная биохимия и биофизика Программа, Мемориальный онкологический центр Слоуна – Кеттеринга, Нью-Йорк, NY 10021.
↵ Кому обращаться с запросами на перепечатку: Университет Рокфеллера, 1230 York Avenue, New York, NY 10021. Электронная почта: Берли rockvax.rockefeller.edu.
Сокращения: pol II, полимераза II; PIC, преинициация сложный; AdMLP, главный поздний промотор аденовируса; cTFIIB, терминал C или ядро TFIIB.
- Авторские права © 1997, Национальная академия наук США
Функциональный анализ сайтов связывания факторов транскрипции в промоторах человека | Genome Biology
Lemon B, Tjian R: Организованный ответ: симфония факторов транскрипции для контроля генов. Genes Dev. 2000, 14: 2551-69. 10.1101 / gad.831000.
PubMed Google Scholar
Батлер Дж., Кадонага Дж .: Основной промотор РНК-полимеразы II: ключевой компонент в регуляции экспрессии генов.Genes Dev. 2002, 16: 2583-2592. 10.1101 / gad.1026202.
PubMed Google Scholar
Hannenhalli S: Моделирование сайтов связывания эукариотических факторов транскрипции и методы интегративного поиска. Биоинформатика. 2008, 24: 1325-31. 10.1093 / биоинформатика / btn198.
PubMed Google Scholar
Чжоу К., Вонг WH: CisModule: новое открытие цис-регуляторных модулей с помощью иерархического моделирования смеси.Proc Natl Acad Sci USA. 2004, 101: 12114-9. 10.1073 / pnas.0402858101.
PubMed PubMed Central Google Scholar
Zhu Z, Shendure J, Church GM: обнаружение функциональных комбинаций транскрипционных факторов в клеточном цикле человека. Genome Res. 2005, 15: 848-55. 10.1101 / гр.3394405.
PubMed PubMed Central Google Scholar
Папе У., Кляйн Х., Вингрон М: Статистическое обнаружение кооперативных факторов транскрипции с регулировкой сходства.Биоинформатика. 2009, 25: 2103-2109. 10.1093 / биоинформатика / btp143.
PubMed PubMed Central Google Scholar
Булык М.Л.: Расчетное предсказание местоположений сайтов связывания транскрипционных факторов. Genome Biol. 2003, 5: 201-10.1186 / GB-2003-5-1-201.
PubMed PubMed Central Google Scholar
Фрит М., Ли М., Вен З .: Cluster-Buster: поиск плотных кластеров мотивов в последовательностях ДНК.Nucleic Acids Res. 2003, 31: 3666-3668. 10.1093 / нар / гкг540.
PubMed PubMed Central Google Scholar
Фрит М., Фу Й, Ю Л, Чен Дж, Хансен У., Вен З .: Обнаружение функциональных мотивов ДНК посредством статистического избыточного представления. Nucleic Acids Res. 2004, 32: 1372-1381. 10.1093 / нар / гх399.
PubMed PubMed Central Google Scholar
Tompa M, Li N, Bailey TL, Church GM, Moor BD, Eskin E, Favorov AV, Frith MC, Fu Y, Kent WJ, Makeev VJ, Mironov AA, Noble WS, Pavesi G, Pesole G , Régnier M, Simonis N, Sinha S, Thijs G, van Helden J, Vandenbogaert M, Weng Z, Workman C, Ye C, Zhu Z: Оценка вычислительных инструментов для открытия сайтов связывания транскрипционных факторов.Nat Biotechnol. 2005, 23: 137-44. 10.1038 / nbt1053.
PubMed Google Scholar
Elnitski L, Jin VX, Farnham PJ, Jones SJM: Определение сайтов связывания факторов транскрипции млекопитающих: обзор вычислительных и экспериментальных методов. Genome Res. 2006, 16: 1455-64. 10.1101 / гр. 4140006.
PubMed Google Scholar
Хокинс Дж., Грант С., Нобл В.С., Бейли Т.Л.: Оценка моделей филогенетических мотивов для прогнозирования сайтов связывания факторов транскрипции.Биоинформатика. 2009, 25: I339-I347. 10.1093 / биоинформатика / btp201.
PubMed PubMed Central Google Scholar
Stormo GD: сайты связывания ДНК: представление и открытие. Биоинформатика. 2000, 16: 16-23. 10.1093 / биоинформатика / 16.1.16.
PubMed Google Scholar
Tronche F, Ringeisen F, Blumenfeld M, Yaniv M, Pontoglio M: Анализ распределения сайтов связывания для тканеспецифического фактора транскрипции в геноме позвоночных.J Mol Biol. 1997, 266: 231-245. 10.1006 / jmbi.1996.0760.
PubMed Google Scholar
Керадпур П., Старк А., Рой С., Келлис М.: Надежное предсказание регуляторных мишеней с использованием 12 геномов Drosophila . Genome Res. 2007, 17: 1919-31. 10.1101 / гр.70.
PubMed PubMed Central Google Scholar
Проект ENCODE Консорциум: Проект ENCODE (Энциклопедия элементов ДНК).Наука. 2004, 306: 636-40.
Google Scholar
Консорциум проекта ENCODE: Идентификация и анализ функциональных элементов в 1% генома человека в рамках пилотного проекта ENCODE. Природа. 2007, 447: 799-816. 10.1038 / природа05874.
Google Scholar
Консорциум проекта ENCODE: Руководство пользователя энциклопедии элементов ДНК (ENCODE). PLoS Biol.2011, 9: e1001046-10.1371 / journal.pbio.1001046.
Google Scholar
Myers RM, Tilly K, Maniatis T: генетический анализ тонкой структуры промотора β -глобина. Наука. 1986, 232: 613-8. 10.1126 / science.3457470.
PubMed Google Scholar
Trinklein N, Aldred S, Saldanha A, Myers R: Идентификация и функциональный анализ промоторов транскрипции человека.Genome Res. 2003, 13: 308-312. 10.1101 / гр.794803.
PubMed PubMed Central Google Scholar
Купер С., Тринклейн Н., Антон Э., Нгуен Л., Майерс Р.: Комплексный анализ структуры и функции транскрипционного промотора в 1% генома человека. Genome Res. 2006, 16: 1-10.
PubMed PubMed Central Google Scholar
Ландолин JM, Johnson DS, Trinklein ND, Aldred SF, Medina C, Shulha H, Weng Z, Myers RM: особенности последовательности, которые определяют функцию промотора человека и тканевую специфичность.Genome Res. 2010, 20: 890-898. 10.1101 / gr.100370.109.
PubMed PubMed Central Google Scholar
Матис В., Кель-Маргулис О.В., Фрике Э., Либих I, Лэнд С., Барре-Дирри А., Рейтер I, Чекменев Д., Круль М., Хорнишер К., Фосс Н., Штегмайер П., Левицки-Потапов Б. , Saxel H, Kel AE, Wingender E: TRANSFAC и его модуль TRANSCompel: регуляция транскрипционных генов у эукариот. Nucleic Acids Res. 2006, 34: D108-10. 10.1093 / нар / gkj143.
PubMed PubMed Central Google Scholar
Санделин А., Алкема В., Энгстром П., Вассерман В., Ленхард Б.: JASPAR: база данных с открытым доступом для профилей связывания эукариотических факторов транскрипции. Nucleic Acids Res. 2004, 32: D91-D94. 10.1093 / нар / гх012.
PubMed PubMed Central Google Scholar
Xie X, Mikkelsen TS, Gnirke A, Lindblad-Toh K, Kellis M, Lander ES: систематическое открытие регуляторных мотивов в консервативных областях генома человека, включая тысячи сайтов инсуляторов CTCF.Proc Natl Acad Sci USA. 2007, 104: 7145-7150. 10.1073 / pnas.0701811104.
PubMed PubMed Central Google Scholar
Филлипс Дж. Э., Корсес В.Г.: CTCF: мастер-ткач генома. Клетка. 2009, 137: 1194-1211. 10.1016 / j.cell.2009.06.001.
PubMed PubMed Central Google Scholar
Хандоко Л., Сюй Х, Ли Джи, Нган Си, Чу Э, Шнапп М, Ли КВХ, Йе С, Пинг ДжЛХ, Мулавади Ф, Вонг Э, Шэн Дж, Чжан И, Пох Т, Чан С.С. , Kunarso G, Shahab A, Bourque G, Cacheux-Rataboul V, Sung WK, Ruan Y, Wei CL: CTCF-опосредованный функциональный взаимодействующий хроматин в плюрипотентных клетках.Нат Жене. 2011, 43: 630-U198. 10,1038 / нг.857.
PubMed PubMed Central Google Scholar
Bell A, West A, Felsenfeld G: Белок CTCF необходим для блокирования энхансера активности инсуляторов позвоночных. Клетка. 1999, 98: 387-396. 10.1016 / S0092-8674 (00) 81967-4.
PubMed Google Scholar
Олссон Р., Ренкавиц Р., Лобаненков В.: CTCF — это уникальный универсальный регулятор транскрипции, связанный с эпигенетикой и болезнью.Тенденции Genet. 2001, 17: 520-527. 10.1016 / S0168-9525 (01) 02366-6.
PubMed Google Scholar
Востров А., Китшке В.: Цинковый белок CTCF связывается с бета-доменом APB промотора предшественника амилоидного бета-белка — свидетельство его роли в активации транскрипции. J Biol Chem. 1997, 272: 33353-33359. 10.1074 / jbc.272.52.33353.
PubMed Google Scholar
Fu Y, Sinha M, Peterson CL, Weng Z: белок, связывающий инсулятор, CTCF позиционирует 20 нуклеосом вокруг своих сайтов связывания в геноме человека. PLoS Genet. 2008, 4: e1000138-10.1371 / journal.pgen.1000138.
PubMed PubMed Central Google Scholar
Филиппова Г., Фагерли С., Кленова Е., Майерс С., Денер Ю., Гудвин Г., Нейман П., Коллинз С., Лобаненков В. Исключительно консервативный репрессор транскрипции, CTCF, использует различные комбинации цинковых пальцев для связывания расходящихся промоторные последовательности онкогенов c- myc птиц и млекопитающих.Mol Cell Biol. 1996, 16: 2802-2813.
PubMed PubMed Central Google Scholar
Ламарко К., Томпсон С., Байерс Б., Уолтон Е., Макнайт С. Идентификация связанных с Ets и notch субъединиц в связывающем белке GA. Наука. 1991, 253: 789-792. 10.1126 / science.1876836.
Google Scholar
Янг З.Ф., Мотт С., Розмарин А.Г .: Фактор транскрипции Ets GABP необходим для развития клеточного цикла.Nat Cell Bio. 2007, 9: 339-U164. 10.1038 / ncb1548.
Google Scholar
Yu S, Zhao DM, Jothi R, Xue HH: Критическая потребность в GABP альфа для нормального развития Т-клеток. J Biol Chem. 2010, 285: 10179-10188. 10.1074 / jbc.M109.088740.
PubMed PubMed Central Google Scholar
Ристевски С., О’Лири Д., Торнелл А., Оуэн М., Кола И., Герцог П. Фактор транскрипции ETS GABP альфа необходим для раннего эмбриогенеза.Mol Cell Biol. 2004, 24: 5844-5849. 10.1128 / MCB.24.13.5844-5849.2004.
PubMed PubMed Central Google Scholar
Yu M, Yang XY, Schmidt T., Chinenov Y, Wang R, Martin ME: GA-связывающие белковые элементы инициатора транскрипции. J Biol Chem. 1997, 272: 29060-29067. 10.1074 / jbc.272.46.29060.
PubMed Google Scholar
Orkin S: GATA-связывающие факторы транскрипции в гемопоэтических клетках.Кровь. 1992, 575-581.
Google Scholar
Omichinski J, Trainor C, Evans T, Gronenborn A, Clore G, Felsenfeld G: небольшой пептид с одним пальцем из эритроидного транскрипционного фактора GATA-1 специфически связывается с ДНК в виде комплекса цинка или железа. . Proc Natl Acad Sci USA. 1993, 90: 1676-1680. 10.1073 / pnas.90.5.1676.
PubMed PubMed Central Google Scholar
Молкентин J: факторы транскрипции, содержащие цинковые пальцы GATA-4, -5 и -6 — повсеместно экспрессируемые регуляторы тканеспецифической экспрессии генов. J Biol Chem. 2000, 275: 38949-38952. 10.1074 / jbc.R000029200.
PubMed Google Scholar
Ohneda K, Yamamoto M: Роли гемопоэтических факторов транскрипции GATA-1 и GATA-2 в развитии клонов эритроцитов. Acta Haematol. 2002, 108: 237-245. 10.1159 / 000065660.
PubMed Google Scholar
Pedone PV, Omichinski J, Nony P, Trainor C, Gronenborn A, Clore G, Felsenfeld G: N-концевые пальцы куриных GATA-2 и GATA-3 представляют собой независимые, специфичные для последовательности ДНК-связывающие домены. EMBO J. 1997, 16: 2874-2882. 10.1093 / emboj / 16.10.2874.
PubMed PubMed Central Google Scholar
Suzuki M, Shimizu R, Yamamoto M: регуляция транскрипции с помощью GATA1 и GATA2 во время эритропоэза.Int J Hematol. 2011, 93: 150-155. 10.1007 / s12185-011-0770-6.
PubMed Google Scholar
Tsai J, Tong Q, Tan G, Chang A, Orkin S, Hotamisligil G: Фактор транскрипции GATA2 регулирует дифференцировку коричневых адипоцитов. EMBO Rep.2005, 6: 879-884. 10.1038 / sj.embor.7400490.
PubMed PubMed Central Google Scholar
Yamamoto M, Ko L, Leonard M, Beug H, Orkin S, Engel J: Активность и тканеспецифическая экспрессия мультигенного семейства транскрипционного фактора NF-E1.Genes Dev. 1990, 4: 1650-1662. 10.1101 / gad.4.10.1650.
PubMed Google Scholar
Эль-Вакил А., Франциус С., Вольф А., Пло-Варет Дж., Нарделли Дж.: Фактор транскрипции GATA2 отрицательно регулирует пролиферацию предшественников нейронов. Разработка. 2006, 133: 2155-2165. 10.1242 / dev.02377.
PubMed Google Scholar
Caramori G, Lim S, Ito K, Tomita K, Oates T, Jazrawi E, Chung K, Barnes P, Adcock I: Экспрессия транскрипционных факторов семейства GATA в Т-клетках, моноцитах и биопсиях бронхов.Eur Resipr J. 2001, 18: 466-473. 10.1183 / 036.01.00040701.
Google Scholar
Shureiqi I, Zuo X, Broaddus R, Wu Y, Guan B, Morris JS, Lippman SM: фактор транскрипции GATA-6 сверхэкспрессируется in vivo и способствует подавлению 15-LOX-1 in vitro у человека. рак толстой кишки. FASEB J. 2007, 21: 743-753. 10.1096 / fj.06-6830com.
PubMed PubMed Central Google Scholar
Невинс Дж .: К пониманию функциональной сложности семейств E2F и ретинобластомы. Рост клеток различается. 1998, 9: 585-593.
PubMed Google Scholar
Тримарчи Дж., Лис Дж .: Соперничество братьев и сестер в семье E2F. Nat Rev Mol Cell Biol. 2002, 3: 11-20.
PubMed Google Scholar
Cam H, Dynlacht B: Новые роли E2F: помимо перехода G1 / S и репликации ДНК.Раковая клетка. 2003, 3: 311-316. 10.1016 / S1535-6108 (03) 00080-1.
PubMed Google Scholar
Attwooll C, Denchi E, Helin K: Семейство E2F: особые функции и пересекающиеся интересы. EMBO J. 2004, 23: 4709-4716. 10.1038 / sj.emboj.7600481.
PubMed PubMed Central Google Scholar
Reimer D, Sadr S, Wiedemair A, Goebel G, Concin N, Hofstetter G, Marth C, Zeimet AG: Экспрессия транскрипционных факторов семейства E2F и ее клиническое значение при раке яичников.Пути передачи сигнала, структура хроматина и механизмы экспрессии генов как терапевтические мишени, Med Univ Innsbruck, Dept Obstet & Gynecol, A-6020 Innsbruck, Austria. 2006, 270-281.
Google Scholar
Рабинович А., Джин В.Х., Рабинович Р., Сюй Х, Фарнхэм П.Дж.: E2F-специфичность связывания in vivo: сравнение согласованных и неконсенсусных сайтов связывания. Genome Res. 2008, 18: 1763-1777. 10.1101 / gr.080622.108.
PubMed PubMed Central Google Scholar
Wells J, Boyd K, Fry C, Bartley S, Farnham P: Специфичность целевого гена для E2F и членов семейства карманных белков в живых клетках. Mol Cell Biol. 2000, 20: 5797-5807. 10.1128 / MCB.20.16.5797-5807.2000.
PubMed PubMed Central Google Scholar
Димова Д., Дайсон Н.: Транскрипционная сеть E2F: старые знакомства с новыми лицами. Онкоген. 2005, 24: 2810-2826. 10.1038 / sj.onc.1208612.
PubMed Google Scholar
Christensen J, Cloos P, Toftegaard U, Klinkenberg D, Bracken A, Trinh E, Heeran M, Di Stefano L, Helin K: характеристика E2F8, нового E2F-подобного репрессора E2F-активированной транскрипции, регулируемого клеточным циклом. Nucleic Acids Res. 2005, 33: 5458-5470. 10.1093 / нар / gki855.
PubMed PubMed Central Google Scholar
Ли Б.К., Бинге А.А., Айер В.Р.: Широкий спектр функций E2F4 в активации и репрессии транскрипции выявлен с помощью полногеномного анализа.Nucleic Acids Res. 2011, 39: 3558-3573. 10.1093 / нар / gkq1313.
PubMed PubMed Central Google Scholar
Helin K, Wu C, Fattaey A, Lees J, Dynlacht B, NGWU C, Harlow E: Гетеродимеризация факторов транскрипции E2F-1 и DP-1 приводит к совместной активации транскрипции —. Genes Dev. 1993, 7: 1850-1861. 10.1101 / gad.7.10.1850.
PubMed Google Scholar
Cao AR, Rabinovich R, Xu M, Xu X, Jin VX, Farnham PJ: Полногеномный анализ мутантных белков фактора транскрипции E2F1 показывает, что N- и C-концевые домены взаимодействия белков не участвуют в нацеливании E2F1 на геном человека . J Biol Chem. 2011, 286: 11985-11996. 10.1074 / jbc.M110.217158.
PubMed PubMed Central Google Scholar
Леви Д.Е., Дарнелл Дж. Э .: Статистика: контроль транскрипции и биологическое воздействие.Nat Rev Mol Cell Biol. 2002, 3: 651-662.
PubMed Google Scholar
Darnell JE: STATs и генная регуляция. Наука. 1997, 277: 1630-1635. 10.1126 / science.277.5332.1630.
PubMed Google Scholar
Bowman T, Garcia R, Turkson J, Jove R: STATs в онкогенезе. Онкоген. 2000, 19: 2474-2488. 10.1038 / sj.onc.1203527.
PubMed Google Scholar
Ян Дж., Старк Г. Р.: Роль нефосфорилированных STAT в передаче сигналов. Клеточные исследования. 2008, 18: 443-451. 10.1038 / кр.2008.41.
PubMed Google Scholar
Horvath CM: белки STAT и транскрипционные ответы на внеклеточные сигналы. Trends Biochem Sci. 2000, 25: 496-502. 10.1016 / S0968-0004 (00) 01624-8.
PubMed Google Scholar
Хартман С., Бертон П., Нат А., Ройс Т., Герштейн М., Вайсман С., Снайдер М.: Глобальные изменения в селекции мишеней STAT и регуляции транскрипции при лечении интерфероном.Genes Dev. 2005, 19: 2953-2968. 10.1101 / гад.1371305.
PubMed PubMed Central Google Scholar
Куреши С., Салдит-Джорджи М., Дарнелл Дж. Тирозин-фосфорилированные Stat1 и Stat2 плюс белок 48 кда все контактируют с ДНК при образовании фактора-3 гена, стимулированного интерфероном. Proc Natl Acad Sci USA. 1995, 92: 3829-3833. 10.1073 / pnas.92.9.3829.
PubMed PubMed Central Google Scholar
Li X, Leung S, Qureshi S, Darnell J, Stark G: Образование гетеродимеров STAT1-STAT2 и их роль в активации транскрипции гена IRF-1 интерфероном-альфа. J Biol Chem. 1996, 271: 5790-5794. 10.1074 / jbc.271.10.5790.
PubMed Google Scholar
Сяо В., Линднер Д., Калваколану Д.: индуцируемый интерфероном мышиный ген p48 (ISGF3 гамма) регулируется протоонкогеном c-myc. Proc Natl Acad Sci USA. 1997, 94: 7227-7232.10.1073 / pnas.94.14.7227.
Google Scholar
Martinez-Moczygemba M, Gutch M, French D, Reich N: отчетливая структура STAT способствует взаимодействию STAT2 с субъединицей p48 стимулированного интерфероном-альфа фактора транскрипции ISGF3. J Biol Chem. 1997, 272: 20070-20076. 10.1074 / jbc.272.32.20070.
PubMed Google Scholar
Ghislain J, Wong T, Nguyen M, Fish E: индуцируемый интерфероном гетеродимер Stat2: Stat1 предпочтительно связывается in vitro с согласованным элементом, обнаруженным в промоторах подгруппы генов, стимулированных интерфероном.J Interferon Cytokine Res. 2001, 21: 379-388. 10.1089 / 10799
50277853.
PubMed Google Scholar
Ши Й, Ли Дж, Галвин К. Все, что вы когда-либо хотели знать об Инь Ян 1. Biochim Biophys Acta Rev Cancer. 1997, 1332: F49-F66. 10.1016 / S0304-419X (96) 00044-3.
Google Scholar
Shi Y, Seto E, Chang L, Shenk T: Репрессия транскрипции с помощью Yy1, человеческого белка, связанного с Gli-Kruppel, и облегчение репрессии с помощью белка E1a аденовируса.Клетка. 1991, 67: 377-388. 10.1016 / 0092-8674 (91)
-6.
PubMed Google Scholar
До Ким Дж, Фолк С., Ким Дж .: Ретропозиция и эволюция ДНК-связывающих мотивов YY1, YY2 и REX1. Nucleic Acids Res. 2007, 35: 3442-3452. 10.1093 / нар / гкм235.
Google Scholar
Lee TC, Shi Y, Schwartz RJ: Смещение BrdUrd-индуцированного YY1 фактором ответа сыворотки активирует транскрипцию скелетного альфа-актина в эмбриональных миобластах.Proc Natl Acad Sci USA. 1992, 89: 9814-9818. 10.1073 / pnas.89.20.9814.
PubMed PubMed Central Google Scholar
A Shrivastava KC: анализ генов, регулируемых многофункциональным регулятором транскрипции Инь Ян-1. Nucleic Acids Res. 1994, 22: 5151-10.1093 / nar / 22.24.5151.
PubMed Google Scholar
Xi H, Yu Y, Fu Y, Foley J, Halees A, Weng Z: Анализ чрезмерно представленных мотивов в основных промоторах человека показывает двойную регуляторную роль YY1.Genome Res. 2007, 17: 798-806. 10.1101 / гр.5754707.
PubMed PubMed Central Google Scholar
Cheng Y, Handwerger S: специфичный для плаценты энхансер гена синцитина человека. Биол Репрод. 2005, 73: 500-509. 10.1095 / биолрепрод.105.039941.
PubMed Google Scholar
Уилбердинг Дж., Кастеллино Ф .: Характеристика мышиного фактора свертывания крови × промотор.Thromb Haemost. 2000, 84: 1031-1038.
PubMed Google Scholar
Collins PJ, Kobayashi Y, Nguyen L, Trinklein ND, Myers RM: фактор транскрипции, связанный с ets , GABP управляет двунаправленной транскрипцией. PLoS Genet. 2007, 3: e208-10.1371 / journal.pgen.0030208.
PubMed PubMed Central Google Scholar
Zhang X, Odom D, Koo S, Conkright M, Canettieri G, Best J, Chen H, Jenner R, Herbolsheimer E, Jacobsen E, Kadam S, Ecker J, Emerson B, Hogenesch J, Unterman T. , Young R, Montminy M: полногеномный анализ занятости связывающего белка элемента цАМФ, фосфорилирования и активации гена-мишени в тканях человека.Proc Natl Acad Sci USA. 2005, 102: 4459-4464. 10.1073 / pnas.0501076102.
PubMed PubMed Central Google Scholar
Баннерт Н., Авотс А., Байер М., Серинг Е, Курт Р. Факторы GA-связывающего белка в сочетании с коактиватором CREB-связывающего белка p300 контролируют индукцию промотора интерлейкина 16 в Т-лимфоцитах. Proc Natl Acad Sci USA. 1999, 96: 1541-1546. 10.1073 / pnas.96.4.1541.
PubMed PubMed Central Google Scholar
De Guzman R, Martinez-Yamout M, Dyson H, Wright P: Взаимодействие домена TAZ1 CREB-связывающего белка с доменом активации CITED2 — регуляция за счет конкуренции между внутренне неструктурированными лигандами за неидентичные сайты связывания. J Biol Chem. 2004, 279: 3042-3049.
PubMed Google Scholar
Honda K, Yanai H, Negishi H, Asagiri M, Sato M, Mizutani T, Shimada N, Ohba Y, Takaoka A, Yoshida N, Taniguchi T: IRF-7 — главный регулятор типа I интерферон-зависимые иммунные ответы.Природа. 2005, 434: 772-777. 10.1038 / природа03464.
PubMed Google Scholar
Odom D, Zizlsperger N, Gordon D, Bell G, Rinaldi N, Murray H, Volkert T., Schreiber J, Rolfe P, Gi ord D, Fraenkel E, Bell G, Young R: Контроль поджелудочной железы и Экспрессия гена печени факторами транскрипции HNF. Наука. 2004, 303: 1378-1381. 10.1126 / science.1089769.
PubMed PubMed Central Google Scholar
Gotea V, Visel A, Westlund JM, Nobrega MA, Pennacchio LA, Ovcharenko I. Гомотипические кластеры сайтов связывания факторов транскрипции являются ключевым компонентом промоторов и энхансеров человека. Genome Res. 2010, 20: 565-577. 10.1101 / gr.104471.109.
PubMed PubMed Central Google Scholar
Ван Дж., Чжуан Дж., Айер С., Линь Икс, Уит Элд Т.В., Гревен М., Пирс Б.Г., Донг Х, Кундаже А, Ченг Й., Рандо О.Дж., Бирни Э., Майерс Р.М., Нобл В.С., Синдер M, Weng Z: Особенности последовательности и структура хроматина вокруг геномных областей, связанных 119 факторами транскрипции человека.Genome Res. 2012, 22: 1798-1812. 10.1101 / гр.139105.112.
PubMed PubMed Central Google Scholar
Martin D, Pantoja C, Minán AF, Valdes-Quezada C, Moltó E, Matesanz F, Bogdanović O, de la Calle-Mustienes E, Domínguez O, Taher L, Furlan-Magaril M, Alcina A, Каньон С., Федетц М., Бласко М.А., Перейра П.С., Овчаренко И., Ресиллас-Тарга Ф., Монтолиу Л., Мансанарес М., Гиго Р., Серрано М., Касарес Ф., Гомес-Скармета Ю. Л.: Распределение CTCF по всему геному у позвоночных определяет эквивалентные участки которые помогают идентифицировать гены, связанные с заболеванием.Nat Struct Mol Biol. 2011, 18: 708-714. 10.1038 / nsmb.2059.
PubMed PubMed Central Google Scholar
Wang G, Wang Y, Teng M, Zhang D, Li L, Liu Y: Преобразователи сигналов и активаторы транскрипции-1 (STAT1) регулируют транскрипцию микроРНК в клетках HeLa, стимулированных интерфероном гамма. PLoS ONE. 2010, 5: e11794-10.1371 / journal.pone.0011794.
PubMed PubMed Central Google Scholar
Siepel A, Pollard KS, Haussler D: Новые методы для определения отбора, зависящего от происхождения. Исследования в области вычислительной молекулярной биологии, Слушания, Том 3909 конспектов лекций по информатике. Под редакцией: Апостолико А., Герра С., Истрайл С., Певзнер П. и Уотерман М. 2006, Heidelberger Platz 3, D-14197 Берлин, Германия: Springer-Verlag Berlin, 190-205.
Google Scholar
Kellis M, Patterson N, Endrizzi M, Birren B, Lander ES: Секвенирование и сравнение видов дрожжей для идентификации генов и регуляторных элементов.Природа. 2003, 423: 241-54. 10.1038 / природа01644.
PubMed Google Scholar
Stark A, Lin MF, Kheradpour P, Pedersen JS, Parts L, Carlson JW, Crosby MA, Rasmussen MD, Roy S, Deoras AN, Ruby JG, Brennecke J, кураторы HF, Project BDG, Hodges E , Hinrichs AS, Caspi A, Paten B, Park SW, Han MV, Maeder ML, Polansky BJ, Robson BE, Aerts S, van Helden J, Hassan B, Gilbert DG, Eastman DA, Rice M, Weir M, Hahn MW, Park Y, Dewey CN, Pachter L, Kent WJ, Haussler D, Lai EC, Bartel DP, Hannon GJ, Kaufman TC, Eisen MB, Clark AG, Smith D, Celniker SE, Gelbart WM, Kellis M: открытие функциональных элементов в 12 Drosophila геномов с использованием эволюционных сигнатур.Природа. 2007, 450: 219-32. 10.1038 / природа06340.
PubMed PubMed Central Google Scholar
Blanchette M, Sinha S: Отделение реальных мотивов от их артефактов. Биоинформатика. 2001, 17 (Приложение 1): S30-8. 10.1093 / биоинформатика / 17.suppl_1.S30.
PubMed Google Scholar
Купер Г., Стоун Е., Асименос Г., Грин Е., Бацоглу С., Сидоу А. Распределение и интенсивность ограничений в геномной последовательности млекопитающих.Genome Res. 2005, 15: 901-913. 10.1101 / гр.3577405.
PubMed PubMed Central Google Scholar
Kasowski M, Grubert F, He el nger C, Hariharan M, Asabere A, Waszak SM, Habegger L, Rozowsky J, Shi M, Urban AE, Hong MY, Karczewski KJ, Huber W, Weissman SM, Герштейн М.Б., Корбель Дж.О., Снайдер М.: Вариации связывания факторов транскрипции среди людей. Наука. 2010, 328: 232-5. 10.1126 / science.1183621.
PubMed PubMed Central Google Scholar
McDaniell R, Lee BK, Song L, Liu Z, Boyle AP, Erdos MR, Scott LJ, Morken MA, Kucera KS, Battenhouse A, Keefe D, Collins FS, Willard HF, Lieb JD, Furey TS, Crawford GE, Iyer VR, Birney E: Наследственные индивидуально-специфические и аллель-специфичные сигнатуры хроматина у людей. Наука. 2010, 328: 235-9. 10.1126 / science.1184655.
PubMed PubMed Central Google Scholar
Zhang Y, Liu T, Meyer CA, Eeckhoute J, Johnson DS, Bernstein BE, Nussbaum C, Myers RM, Brown M, Li W, Liu XS: модельный анализ ChIP-Seq (MACS) .Genome Biol. 2008, 9: R137-10.1186 / gb-2008-9-9-r137.
PubMed PubMed Central Google Scholar
Фрит М.: ПОССУМ: обнаружение цис-элементов в последовательностях ДНК. [http://biowulf.bu.edu/MotifViz/]
Fu Y, Weng Z: Улучшение матриц TRANSFAC с использованием множественного локального выравнивания последовательностей сайтов связывания факторов транскрипции. Геномная информатика Международная конференция по геномной информатике. 2005, 16: 68-72.
PubMed Google Scholar
Rosenbloom KR, Dreszer TR, Pheasant M, Barber GP, Meyer LR, Pohl A, Raney BJ, Wang T, Hinrichs AS, Zweig AS, Fujita PA, Learned K, Rhead B, Smith KE, Kuhn RM , Карольчик Д., Хаусслер Д., Кент В. Дж.: ЗАКОДИРУЙТЕ данные полного генома в браузере генома UCSC. Nucleic Acids Res. 2010, 38: D620-D625. 10.1093 / нар / gkp961.
PubMed PubMed Central Google Scholar
Zheng L, Baumann U, Reymond J: эффективный одностадийный сайт-направленный и сайт-насыщенный протокол мутагенеза. Nucleic Acids Res. 2004, 32: e115-10.1093 / nar / gnh210.
PubMed PubMed Central Google Scholar
Schneider T, Stormo G, Gold L, Ehrenfeucht A: информация о сайтах связывания в нуклеотидных последовательностях. J Mol Biol. 1986, 188: 415-431. 10.1016 / 0022-2836 (86)-8.
PubMed Google Scholar
Лин Дж., Коллинз П., Тринкляйн Н., Фу Y, Си Х, Майерс Р., Вен З .: Связывание факторов транскрипции и модифицированные гистоны в двунаправленных промоторах человека. Genome Res. 2007, 17: 818-827. 10.1101 / гр.5623407.
PubMed PubMed Central Google Scholar
Прюитт К., Татусова Т., Маглотт Д.: Контрольные последовательности NCBI (RefSeq): тщательно подобранная база данных неизбыточных последовательностей геномов, транскриптов и белков. Nucleic Acids Res. 2007, 35: Д61-65.10.1093 / нар / gkl842.
PubMed PubMed Central Google Scholar
Хсу Ф., Кент В., Клоусон Х., Кун Р., Диханс М., Хаусслер Д.: известные гены UCSC. Биоинформатика. 2006, 22: 1036-1046. 10.1093 / биоинформатика / btl048.
PubMed Google Scholar
Харроу Дж., Деноуд Ф, Франкиш А., Реймонд А., Чен К. К., Краст Дж., Лагард Дж., Гилберт Дж. Г., Стори Р., Сварбрек Д., Россье С., Укла С., Хаббард Т., Антонаракис С. Е., Гиго Р. : GENCODE: создание справочной аннотации для ENCODE.Genome Biol. 2006, 7 (Дополнение 1): S4 1-9.
Google Scholar
McLean CY, Bristor D, Hiller M, Clarke SL, Schaar BT, Lowe CB, Wenger AM, Bejerano G: GREAT улучшает функциональную интерпретацию цис-регуляторных регионов. Nat Biotechnol. 2010, 28: 495-501. 10.1038 / nbt.1630.
PubMed Google Scholar
Bowie ML, Troch MM, Delrow J, Dietze EC, Bean GR, Ibarra C, Pandiyan G, Seewaldt VL: Интерфероновый регуляторный фактор-1 регулирует апоптоз, опосредованный восстановленным внеклеточным матриксом (rECM) в эпителиальных клетках молочной железы человека .Онкоген. 2007, 26: 2017-2026. 10.1038 / sj.onc.1210013.
PubMed Google Scholar
Grandvaux N, Servant MJ, tenOever B, Sen GC, Balachandran S, Barber GN, Lin R, Hiscott1 J: Транскрипционное профилирование генов-мишеней регулирующего фактора интерферона 3: прямое участие в регуляции генов, стимулированных интерфероном . J Virol. 2002, 76: 5532-5539. 10.1128 / JVI.76.11.5532-5539.2002.
PubMed PubMed Central Google Scholar
Промоутеры — отраслевой сайт OBRA
Новый промоутер?
Вы промоутер, который хочет устроить гонку OBRA? Есть вопросы? Ваш первый шаг — прочитать руководство промоутера.
Оборудование
Тони Кич — наш эксперт по оборудованию. С любыми вопросами обращайтесь по электронной почте [email protected] и ознакомьтесь с нашей формой запроса оборудования слева. Обязательно сообщите Тони, если какое-либо оборудование неисправно или не работает, чтобы мы могли вывести его из ротации.
Отправка события
После того, как вы определились, что собираетесь проводить мероприятие, отправьте запрос даты, используя эту форму.
Заявка на участие в общественном мероприятии, не относящемся к расе
Пожалуйста, используйте форму здесь, чтобы подать заявку на участие в велоспорте, не относящемся к гонкам.
Маркетинговые услуги
Мы наняли нового координатора по маркетингу Джоэла Флетчера, чтобы он помог OBRA и промоутерам с маркетингом. С ним можно связаться по адресу [email protected]. У нас также есть множество инструментов, которые помогут вам продвигать свою продукцию и получить обратную связь. У нас есть список адресов электронной почты, содержащий более 17 000 адресов электронной почты участников гонки. У нас также есть подписка на SurveyMonkey. Если вы хотите провести опрос или получить доступ к списку адресов электронной почты, сообщите нам об этом.
Онлайн-регистрация
Каждый промоутер может использовать собственную службу онлайн-регистрации.Однако у OBRA есть собственная служба онлайн-регистрации. Для получения дополнительной информации посетите страницу с информацией о онлайн-регистрации на главном сайте OBRA.
OBRA Первая помощь
Когда промоутеры выбирают использование первой помощи OBRA (доступно в некоторых регионах) — вы получаете программу, которая помогает гонщикам, когда гонщик получает травму. Они промывают и перевязывают ваши раны. Программа взимает плату только за время оказания первой помощи. Все материалы предоставляются OBRA без дополнительной оплаты промоутеру или гонщику.Свяжитесь с [email protected] для получения подробной информации.
Форумы
Мы создали форум для промоутеров для обсуждения вопросов. Вам потребуется разрешение на доступ к нашим форумам промоутеров.
Оплата гонок
Щелкните здесь, чтобы перейти в наш промышленный магазин.
Промоутеров — parts.igem.org
Символ реестра для промотора показан над типичной последовательностью для бактериального промотора. Закрашенные бледно-лиловым цветом прямоугольники указывают на две наиболее консервативные области бактериального промотора и расположены на -10 и -35 оснований от сайта начала транскрипции (заштрихованы зеленым).Между сайтами -10 и -35 в среднем 17 п.н. и 7 п.н. между сайтом -10 и сайтом начала транскрипции pribnow harley lisser1 lisser2 .Промотор — это последовательность ДНК, которая может задействовать транскрипционный аппарат и приводить к транскрипции следующей последовательности ДНК. Конкретная последовательность промотора определяет силу промотора (сильный промотор приводит к высокой скорости инициации транскрипции).
В дополнение к последовательностям, которые «стимулируют» транскрипцию, промотор может включать дополнительные последовательности, известные как операторы, которые контролируют силу промотора.Например, промотор может включать сайт связывания для белка, который привлекает или препятствует связыванию RNAP с промотором. Присутствие или отсутствие белка повлияет на силу промотора. Такой промотор известен как регулируемый промотор.
Описание ввода / вывода функции промоутера
Иногда мы игнорируем детали того, как работает промоутер, и думаем о промоутере как об устройстве, которое преобразует входные данные в выходные. Это можно сделать при разработке многокомпонентной системы, включающей промоторы, активность которых должна регулироваться другими видами в системе.Промотор можно рассматривать как устройство, которое выводит определенное количество транскрибирующих РНК-полимераз в единицу времени. У промоутеров может быть разное количество входов. Конституционный промотор не имеет входов. Технически, даже конститутивный промотор имеет входные данные, такие как уровень свободной РНК-полимеразы, но мы часто предполагаем, что уровни свободной РНК-полимеразы либо неизменны, либо никогда не являются ограничивающим фактором в инициации транскрипции. Уровень репрессора, который негативно регулирует промотор, является входом для промотора.